")
")
| Issue |
Med Sci (Paris)
Volume 34, Number 2, Février 2018
|
|
|---|---|---|
| Page(s) | 161 - 165 | |
| Section | M/S Revues | |
| DOI | https://doi.org/10.1051/medsci/20183402014 | |
| Published online | 16 February 2018 | |
Séquençage de l’ADN par nanopores
Résultats et perspectives
DNA sequencing by nanopores: achievements and prospects
Univ Lyon, Ens de Lyon, Univ Claude Bernard, CNRS, Laboratoire de Physique, 46, allée d’Italie, F-69342 Lyon, France
*
This email address is being protected from spambots. You need JavaScript enabled to view it.
Résumé
Après des années de développement, l’utilisation du nanopore comme sonde pour séquencer les molécules d’ADN est maintenant une possibilité viable et prometteuse. La détection d’une seule paire de bases lors du transport de l’ADN permet d’enregistrer de très longs fragments de polynucléotides, avec une parallélisation et des vitesses élevées. Dans cette revue, les méthodologies actuelles fondées sur la détection électrique et les nanopores biologiques seront présentées de même que les nouvelles méthodes utilisant des nanopores à l’état solide, ou la détection optique.
Abstract
After years of development, the use of nanopore as a sensor to sequence DNA molecules is now a viable and promising possibility. Single base pair detection during DNA transport enables to record ultra-long threads with high parallelization and rates. I will present in this review the current methodologies based on electrical detection and biological nanopores and the new methods based on solid state nanopores and optical detection.
© 2018 médecine/sciences – Inserm

Vignette (Photo © Inserm - Patrick Delapierre).
Séquençage de l’ADN : méthodes commerciales actuelles
Les techniques commerciales actuelles (Illumina, Ion Torrent, Pacific Biosciences) de séquençage de l’ADN sont fondées sur un signal obtenu lors de l’amplification ou la synthèse de molécules individuelles [1-3]. Les techniques les plus couramment utilisées (Illumina, Ion Torrent) se fondent sur l’amplification de l’ADN par PCR (polymerase chain reaction) pour amplifier le brin d’ADN à séquencer, sur une microbille ou une surface fonctionnalisée. Dans le cas d’Illumina, le résultat de cette étape est ensuite séquencé optiquement par synthèse. L’intérêt de cette méthode est sa très grande capacité d’analyse en parallèle (parallélisation) (1 à 100 x 109 lectures [reads] obtenus en 10 à 40 h). Son défaut principal est la longueur des fragments obtenus (< 300 bases) qui nécessite une étape complexe d’alignement des fragments séquencés. Elle est actuellement largement utilisée pour le re-séquençage de génomes entiers, le séquençage de novo, et pour les séquençages ciblés. Une autre technique, développée par Ion Torrent [4, 5], est fondée sur la mesure du pH. En pratique, il s’agit également d’un séquençage par synthèse qui détecte sur un transistor à effet de champ, l’intégration d’une paire de bases par la production d’un ion H+. Elle permet, elle aussi, de réaliser un séquençage à partir de molécules uniques, et de lire des séquences plus longues (400 bases) que la technique précédente, mais avec une parallélisation plus faible (1 à 100 x 106 lectures obtenues en 4 h). Elle est surtout utilisée pour des séquençages ciblés. Enfin, la technologie développée par Pacific Biosciences utilise l’amplification optique du signal d’intégration d’une base par l’ADN polymérase afin de réaliser une détection directe à l’échelle de la molécule unique (voir ci- dessous). Elle détecte des fragments longs (supérieurs à 10 kilobases [kb]) mais avec une faible parallélisation (365 000 lectures obtenues par cellule en 10 h). Elle est utilisée pour le re-séquençage de génomes entiers, le séquençage de novo, et pour le séquençage ciblé.
La difficulté d’obtenir des fragments séquencés de grandes longueurs, et l’intérêt d’une lecture en temps réel, a conduit ces dernières années à des recherches sur l’utilisation de nanopores biologiques, ou artificiels, comme outils de séquençage de l’ADN.
Les nanopores utilisés peuvent être soit des protéines formant naturellement des pores au sein de bicouches lipidiques, soit des ouvertures nanométriques réalisées par gravure dans des membranes solides ultrafines (par exemple, en nitrure de silicium).
Séquençage par nanopore : mesure et forçage électrique
L’idée d’utiliser un nanopore pour séquencer l’ADN repose sur le fait qu’il peut être considéré comme une sonde locale que l’ADN serait forcé de traverser de manière séquentielle pour pouvoir être lu base par base. Les premières avancées dans le domaine ont été réalisées par Kasianowicz, Branton et Deamer [6], qui ont montré qu’une différence de potentiel appliquée de part et d’autre d’un nanopore biologique (en l’occurrence, l’alpha-hémolysine isolée de la bactérie Staphylococcus aureus) permettait de forcer des macromolécules (ADN et ARN simple brin) à transiter par cette nano-cavité. En pratique, c’est le confinement des lignes des champs électriques (Figure 1, lignes continues bleues), à l’intérieur du pore, qui permet d’appliquer les forces suffisantes pour aligner et induire la translocation des molécules de tailles bien plus importantes que celle du pore. La détection du passage de ces molécules est réalisée par la mesure du courant ionique traversant le pore : un pore vide correspond à un courant élevé; La présence d’une macromolécule, telle que l’ADN, réduit le flux d’ions au travers du pore, et permet de détecter le passage d’une molécule individuelle (Figure 1, encart haut).
Cette méthode a, dès lors, été appliquée à la détection d’autres macromolécules (le polyéthylène glycol [PEG] [7] ou des protéines [8, 9]), à la caractérisation de la distribution de masse d’un polymère [7], mais aussi à l’étude de la conformation de macromolécules (structures secondaires et tertiaires [8, 10], nœuds [11], association spécifique de protéines [12]).
Pour pouvoir identifier chacune des paires de base de la molécule transportée dans le nanopore, il a fallu résoudre de nombreuses difficultés techniques et améliorer fortement les performances du système. Pour réduire le bruit de mesure, et augmenter la résolution temporelle (détection par unité de temps), la surface des membranes a été réduite (permettant ainsi une diminution du temps de relaxation électrique du support). Le dispositif expérimental et, en particulier l’électrode de mesure, a été directement intégré au sein d’une puce comportant l’étage de préamplification (ce qui a permis une réduction du bruit électrique). La vitesse de translocation trop élevée, par advection électrique1 à travers le nanopore biologique, a été diminuée par le positionnement, en amont du pore, d’enzymes capables de se lier et de transloquer l’ADN simple brin processivement (c’est-à-dire sans se détacher avant l’extrémité du fragment) comme les hélicases ou l’ADN polymérase. Le pore biologique lui-même a été modifié génétiquement afin d’augmenter le temps d’interaction entre l’ADN et les acides aminés présents dans la partie la plus étroite du pore (ou constriction), par mutagénèse dirigée réalisée en particulier sur les gènes codant le pore CsgG de la bactérie Escherichia coli [13]2 La membrane lipidique a également été remplacée par une membrane polymérique, plus stable dans le temps. Enfin, pour obtenir la capacité de discerner chacune des paires de bases, la réduction de courant ionique induit par chacune d’elle, ainsi que son temps de résidence au sein du nanopore, ont été pris en compte et utilisés. Les modèles d’inférences statistiques (prédiction probabiliste) actuels décomposent le signal en unités glissantes (ou fenêtres mobiles) de 5 bases (5-mers) reproduisant la longueur du fragment piégé dans la constriction (par des approches statistiques [14] et bioinformatiques [15, 16]).
À l’issue de ce processus d’amélioration, cette technique, commercialisée par Oxford Nanopore Technologies [17, 48] ( ) et fondée sur des pores biologiques et la détection électrique, permet d’extraire la séquence de longs fragments individuels de 10 à 1 000 kb, ce qui autorise la compensation par redondance d’un taux d’erreur total élevé. Le taux d’erreur brut total est, dans le meilleur des cas, de 7,5 % pour la lecture d’une base individuelle chez E. coli, en comptant erreurs, insertions et délétions [18]. En utilisant des méthodes par consensus (redondance), la précision progresse jusqu’à 99,5 %, pour une couverture de 30 fois la longueur du génome chez la bactérie E. coli [19]. La vitesse de lecture est, par nanopore, de 250 à 450 bases par seconde [13, 18]. La parallélisation est plus faible que celle obtenue par les techniques d’amplification actuelles (512 à 144 000 pores en parallèle, selon le modèle, et jusqu’à 400 000 lectures par puce de 512 pores [13]). Le dispositif a également un coût, d’un millier d’euros, mais un encombrement qui est similaire à un téléphone portable (pour les systèmes à 512 pores). Cette méthode a permis récemment de réaliser le séquençage, directement sur site, d’espèces de bactéries sauvages [20, 21], de détecter et d’identifier des virus in situ (virus de la dengue [22], parvovirus [23], virus Ebola [24]). Elle a été utilisée pour la détection d’éléments transposables [25], de méthylation de l’ADN [26], et de mutations précoces liées à la maladie d’Alzheimer [27]. Grâce à des algorithmes fondés sur le maximum de vraisemblance, elle a aussi révélé des isoformes de récepteurs dans des lymphocytes B [28]. Des variants structuraux et des mutations dans le cancer du pancréas et dans la leucémie [29, 30] ont pu aussi être détectés dans des études dans lesquelles la méthode s’est montrée complémentaire de celles existantes. Elle a également été testée avec succès pour la détection prénatale d’aneuploïdies dans le sang maternel [31]. Enfin, le séquençage de novo d’un génome humain a récemment été rapporté sur le site de preprint BioRxiv [13]. Des protocoles ont été développés pour augmenter la reproductibilité de ces mesures [32]. Des comparaisons avec les autres techniques de séquençage couramment utilisées ont pu montrer que cette méthode est aujourd’hui compétitive pour le séquençage de novo et l’analyse de variants [17, 18, 33, 34].
) et fondée sur des pores biologiques et la détection électrique, permet d’extraire la séquence de longs fragments individuels de 10 à 1 000 kb, ce qui autorise la compensation par redondance d’un taux d’erreur total élevé. Le taux d’erreur brut total est, dans le meilleur des cas, de 7,5 % pour la lecture d’une base individuelle chez E. coli, en comptant erreurs, insertions et délétions [18]. En utilisant des méthodes par consensus (redondance), la précision progresse jusqu’à 99,5 %, pour une couverture de 30 fois la longueur du génome chez la bactérie E. coli [19]. La vitesse de lecture est, par nanopore, de 250 à 450 bases par seconde [13, 18]. La parallélisation est plus faible que celle obtenue par les techniques d’amplification actuelles (512 à 144 000 pores en parallèle, selon le modèle, et jusqu’à 400 000 lectures par puce de 512 pores [13]). Le dispositif a également un coût, d’un millier d’euros, mais un encombrement qui est similaire à un téléphone portable (pour les systèmes à 512 pores). Cette méthode a permis récemment de réaliser le séquençage, directement sur site, d’espèces de bactéries sauvages [20, 21], de détecter et d’identifier des virus in situ (virus de la dengue [22], parvovirus [23], virus Ebola [24]). Elle a été utilisée pour la détection d’éléments transposables [25], de méthylation de l’ADN [26], et de mutations précoces liées à la maladie d’Alzheimer [27]. Grâce à des algorithmes fondés sur le maximum de vraisemblance, elle a aussi révélé des isoformes de récepteurs dans des lymphocytes B [28]. Des variants structuraux et des mutations dans le cancer du pancréas et dans la leucémie [29, 30] ont pu aussi être détectés dans des études dans lesquelles la méthode s’est montrée complémentaire de celles existantes. Elle a également été testée avec succès pour la détection prénatale d’aneuploïdies dans le sang maternel [31]. Enfin, le séquençage de novo d’un génome humain a récemment été rapporté sur le site de preprint BioRxiv [13]. Des protocoles ont été développés pour augmenter la reproductibilité de ces mesures [32]. Des comparaisons avec les autres techniques de séquençage couramment utilisées ont pu montrer que cette méthode est aujourd’hui compétitive pour le séquençage de novo et l’analyse de variants [17, 18, 33, 34].
()Voir la Chronique génomique de B. Jordan, m/s n° 8-9, août-septembre 2017, page 801
Aujourd’hui, les recherches académiques et industrielles cherchent à étendre le principe de séquençage par nanopore à d’autres biomolécules, en particulier aux polypeptides [35]. Ce problème est cependant notablement plus complexe en raison du grand nombre d’acides aminés, de leurs propriétés physico-chimiques très variables (hydropho- bicité, charge, taille), ainsi que de la structure tridimensionnelle des protéines.
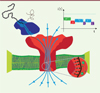 |
Figure 1. Séquençage par nanopore biologique. L’ADN à séquencer est capté par une enzyme (hélicase ou ADN polymérase, en bleu) qui extrude l’ADN simple brin à une vitesse réduite. Le nanopore lui-même (en rouge) constitue une ouverture dans la membrane lipidique (en vert) dans laquelle le champ électrique (flèches bleues) force le passage de l’ADN simple brin (en marron). Ce dernier est ralenti au passage par la constriction du nanopore (encart du bas) et permet l’identification des bases individuelles par mesure de l’intensité du courant électrique traversant le pore à un instant donné (encart du haut). Le trait pointillé bleu représente le courant à vide du nanopore. |
Séquençage par nanopore : pores à l’état solide et détection optique
Les méthodes développées actuellement par plusieurs équipes de recherche tendent à utiliser les nanopores à l’état solide. Ces nanopores sont obtenus par perçage de membranes de nitrure de silicium ou de carbure de silicium ultrafines (de 1 à 50 nm). La cavité est réalisée par un faisceau d’électrons focalisés, ou un faisceau d’ions (gallium, hélium). Le principal intérêt de ces structures est de présenter une géométrie (diamètre, forme du canal) et une organisation (pore unique, réseau de pore) définies lors du perçage [36]. En outre, il à été récemment montré par les groupes de Radenovic [37], Dekker [38, 39] et Drndic [40], que ces pores pouvaient être réalisés dans des membranes bidimensionnelles, c’est-à-dire dans des matériaux ne comportant qu’une seule couche atomique (Figure 2). La résolution temporelle et spatiale de la mesure électrique étant inversement proportionnelle à l’épaisseur de la membrane, ce type de membrane permet d’améliorer grandement la vitesse de lecture des bases et l’identification des nucléotides individuels [37]. Un problème récurrent des nanopores artificiels repose, néanmoins, sur la difficulté de leur conservation et sur leur robustesse d’utilisation; la raison principale étant la réorganisation des atomes au bord du pore, et l’adsorption non spécifique de biomolécules sur ses parois. Les derniers résultats obtenus sur le sujet, proposent d’utiliser des pores percés avec une plus grande énergie de perçage, via un faisceau d’ions (focalized ion beam, FIB) [41] et de fonctionnaliser la surface par des lipides (liposomes) afin de limiter les interactions [42]. Une autre piste de recherche actuelle est d’utiliser des solutions de liquides ioniques pour remplacer les solutions salines [37]. Ces solutions ont une forte conductivité et permettent d’augmenter sensiblement le rapport signal sur bruit de la méthode. Elles sont aussi très visqueuses et favorisent le ralentissement lors du passage de biomolécules au sein du pore.
La détection optique des nucléotides, lors du séquençage par nanopore, est aujourd’hui en plein développement afin de dépasser les limites inhérentes à leur détection électrique. L’utilisation de marquage spécifique de chaque base permet, notamment, de multiplexer l’information et de faciliter l’identification des bases. Les premiers résultats significatifs sur ce sujet ont été obtenus dans le groupe de Meller, qui a montré que la conversion chimique locale de la séquence (remplacement de chaque nucléotide par un groupe de nucléotides prédéfini) permettait de lire optiquement la séquence de l’ADN en cours de translocation dans le nanopore [43]. Les recherches actuelles utilisent un effet optique qui a été découvert par Levene et al., appelé zero-mode waveguide [44] : si un faisceau laser est focalisé sur un puits métallique de quelques nanomètres de diamètre, la lumière ne peut être transmise au sein du pore (limite de diffraction) mais une onde évanescente est excitée localement, à l’entrée du nanopore. En plus de la localisation du champ électrique, le guide d’onde induit un renforcement du champ électrique au niveau de l’entrée du pore. Cet effet a été tout d’abord utilisé pour des nanopuits comportant une ADN polymérase. Dans ce cas, il est possible de suivre l’intégration de bases fluorescentes et ainsi la synthèse du fragment d’ADN. Ces résultats ont conduit à la constitution d’une méthode prometteuse de séquençage, portée par la société Pacific Biosciences [3]. Cependant, pour pouvoir s’affranchir de l’étape de synthèse par l’ADN polymérase, qui limite la vitesse de lecture, de nouvelles recherches ont adapté le principe du zero-mode waveguide à des nanopores [45-47] (Figure 3). Dans ce cas, la lecture ne s’effectue pas par synthèse, mais de manière continue et processive, lors du transport de la molécule comme lors de la détection électrique. Chaque base est marquée avec une molécule fluorescente, le passage devant l’onde évanescente permet d’éclairer spécifiquement cette base et de reconstruire la séquence du fragment.
L’un des intérêts importants des méthodes optiques pour le séquençage par nanopore, est la possibilité de réaliser la mesure avec une très importante parallélisation. Elles permettent aussi de découpler de la détection le forçage de l’ADN au sein du pore. Il est alors possible d’utiliser des solutions salines moins concentrées, plus compatibles avec de potentielles modifications épigénétiques de l’ADN, ou avec des peptides.
 |
Figure 2. Séquençage par nanopore artificiel. L’ADN simple brin (ADNsb en marron) est entraîné par le champ électrique (flèches bleues) à travers un nanopore formé par perçage (faisceau d’électrons issus d’un microscope électronique à transmission [TEM] ou faisceau d’ions [FIB]) dans (A) une membrane ultrafine en nitrure de silicium (SiN), ou (B) un matériau bidimensionnel (disulfure de molybdène [MoS2], graphène, nitrure de bore [BN]). L’identification des bases se fait par la mesure du courant ionique traversant le nanopore. |
 |
Figure 3. Séquençage par nanopore artificiel et détection optique. L’ADN est entraîné par un champ électrique ou un flux hydrodynamique à l’intérieur d’un nanopore artificiel. Lorsque qu’une portion de l’ADN atteint l’extrémité du pore, elle est illuminée par un champ électromagnétique évanescent (en vert) qui induit la fluorescence séquentielle de chacune des paires de bases. Les bases sont alors identifiées par un marqueur fluorescent spécifique. ADNsb : ADN simple brin. |
Conclusion
Les techniques de séquençage par nanopore ont aujourd’hui atteint une maturité technique suffisante pour pouvoir être comparées aux techniques déjà existantes. La nécessité d’augmenter la parallélisation et de diminuer le taux d’erreur de la mesure est toutefois un objectif impératif pour pouvoir les remplacer à long terme et dépasser leur statut actuel de méthode complémentaire dans certaines applications. Les atouts majeurs de cette méthode sont sa capacité à réaliser la lecture de longs fragments, ce qui réduit fortement le travail post-séquençage d’alignement, et aussi la possibilité de construire des dispositifs expérimentaux à la fois moins coûteux et portatifs. Elles ouvrent aussi la voie à des méthodes de séquençage peptidique utilisant le même principe.
Liens d’intérêt
L’auteur déclare n’avoir aucun lien d’intérêt concernant les données publiées dans cet article.
Entraînement de la molécule sous l’effet des forces électriques.
CsgG code une lipoprotéine, composant du système de sécrétion de type VIII de la bactérie et qui forme un canal dans la paroi bactérienne.
Références
- Metzker ML. Sequencing technologies: the next generation. Nat Rev Genet 2010; 11 : 31-46. [Google Scholar]
- Levy SE, Myers RM. Advancements in next-generation sequencing. Annu Rev Genomics Hum Genet 2016; 17 : 95-115. [CrossRef] [Google Scholar]
- Nakano K, Shiroma A, Shimoji M, et al. Advantages of genome sequencing by long-read sequencer using SMRT technology in medical area. Hum Cell 2017; 30 : 149-61. [Google Scholar]
- Merriman B, Torrent I, Rothberg JM. Progress in Ion Torrent semiconductor chip based sequencing. Electrophoresis 2012; 33 : 3397-417. [Google Scholar]
- Rothberg JM, Hinz W, Rearick TM, et al. An integrated semiconductor device enabling non-optical genome sequencing. Nature 2011; 475 : 348-52. [Google Scholar]
- Kasianowicz JJ, Brandin E, Branton D, et al. Characterization of individual polynucleotide molecules using a membrane channel. Proc Natl Acad Sci USA 1996; 93 : 13770-3. [CrossRef] [PubMed] [Google Scholar]
- Robertson JWF, Rodrigues CG, Stanford VM, et al. Single-molecule mass spectrometry in solution using a solitary nanopore. Proc Natl Acad Sci USA 2007; 104 : 8207-11. [CrossRef] [Google Scholar]
- Oukhaled G, Mathé J, Biance A-L, et al. Unfolding of proteins and long transient conformations detected by single nanopore recording. Phys Rev Lett 2007; 98 : 158101. [Google Scholar]
- Oukhaled A, Cressiot B, Bacri L, et al. Dynamics of completely unfolded and native proteins through solid-state nanopores as a function of electric driving force. ACS Nano 2011; 5 : 3628-38. [Google Scholar]
- Mathé J, Visram H, Viasnoff V, et al. Nanopore unzipping of individual DNA hairpin molecules. Biophys J 2004; 87 : 3205-12. [Google Scholar]
- Plesa C, Verschueren D, Pud S, et al. Direct observation of DNA knots using a solid-state nanopore. Nat Nanotechnol 2016; 11 : 1093-7. [Google Scholar]
- Kowalczyk SW, Hall AR, Dekker C. Detection of local protein structures along DNA using solidstate nanopores. Nano Lett 2010; 10 : 324-8. [Google Scholar]
- Jain M, Koren S, Quick J, et al. Nanopore sequencing and assembly of a human genome with ultralong reads. 2017. bioRxiv 128835; doi : https : //doi.org/10.1101/128835. [Google Scholar]
- Goodwin S, Gurtowski J, Ethe-Sayers S, et al. Oxford Nanopore sequencing, hybrid error correction, and de novo assembly of a eukaryotic genome. Genome Res 2015; 25 : 1750-6. [Google Scholar]
- Jain M, Fiddes IT, Miga KH, et al. Improved data analysis for the MinION nanopore sequencer. Nat Methods 2015; 12 : 351-6. [Google Scholar]
- Loman NJ, Quinlan AR. Poretools: a toolkit for analyzing nanopore sequence data. Bioinformatics 2014; 30 : 3399-401. [Google Scholar]
- Jain M, Olsen HE, Paten B, et al. The Oxford Nanopore MinION: delivery of nanopore sequencing to the genomics community. Genome Biol 2016; 17 : 239. [Google Scholar]
- Jain M, Tyson JR, Loose M, et al. MinION analysis and reference consortium: Phase 2 data release and analysis of R9.0 chemistry. F1000Research 2017; 6 : 760. [Google Scholar]
- Loman NJ, Quick J, Simpson JT. A complete bacterial genome assembled de novo using only nanopore sequencing data. Nat Methods 2015; 12 : 733-5. [Google Scholar]
- Ludden C, Reuter S, Judge K, et al. Sharing of carbapenemase-encoding plasmids between Enterobacteriaceae in UK sewage uncovered by MinION sequencing. Microb Genom 2017; 3 : e000114. [Google Scholar]
- Schmidt K, Mwaigwisya S, Crossman LC, et al. Identification of bacterial pathogens and antimicrobial resistance directly from clinical urines by nanopore-based metagenomic sequencing. J Antimicrob Chemother 2017; 72 : 104-14. [CrossRef] [PubMed] [Google Scholar]
- Yamagishi J, Runtuwene LR, Hayashida K, et al. Serotyping dengue virus with isothermal amplification and a portable sequencer. Sci Rep 2017; 7 : 3510. [Google Scholar]
- Batovska J, Lynch SE, Rodoni BC, et al. Metagenomic arbovirus detection using MinION nanopore sequencing. J Virol Methods 2017; 249 : 79-84. [Google Scholar]
- Quick J, Loman NJ, Duraffour S, et al. Real-time, portable genome sequencing for Ebola surveillance. Nature 2016; 530 : 228-32. [Google Scholar]
- Debladis E, Llauro C, Carpentier MC, et al. Detection of active transposable elements in Arabidopsis thaliana using Oxford Nanopore Sequencing technology. BMC Genomics 2017; 18 : 537. [Google Scholar]
- Simpson JT, Workman RE, Zuzarte PC, et al. Detecting DNA cytosine methylation using nanopore sequencing. Nat Methods 2017; 14 : 407-10. [Google Scholar]
- Roeck A De, Bossche T Van den, Zee J van der, et al. Deleterious ABCA7 mutations and transcript rescue mechanisms in early onset Alzheimer’s disease. Acta Neuropathol 2017; 134 : 475-87. [Google Scholar]
- Byrne A, Beaudin AE, Olsen HE, et al. Nanopore long-read RNAseq reveals widespread transcriptional variation among the surface receptors of individual B cells. Nat Commun 2017; 8 : 16027. [Google Scholar]
- Norris AL, Workman RE, Fan Y, et al. Nanopore sequencing detects structural variants in cancer. Cancer Biol Ther 2016; 17 : 246-53. [Google Scholar]
- Minervini CF, Cumbo C, Orsini P, et al. Mutational analysis in BCR - ABL1 positive leukemia by deep sequencing based on nanopore MinION technology. Exp Mol Pathol 2017; 103 : 33-7. [Google Scholar]
- Cheng SH, Jiang P, Sun K, et al. Noninvasive prenatal testing by nanopore sequencing of maternal plasma DNA: feasibility assessment. Clin Chem 2015; 61 : 1305-6. [Google Scholar]
- Goodwin S, Wappel R, McCombie WR. 1D genome sequencing on the Oxford nanopore MinION. Curr Protoc Hum Genet 2017; 94 : 18.11.1-18.11.14. [Google Scholar]
- Judge K, Harris SR, Reuter S, et al. Early insights into the potential of the Oxford Nanopore MinION for the detection of antimicrobial resistance genes. J Antimicrob Chemother 2015; 70 : 2775-8. [CrossRef] [PubMed] [Google Scholar]
- Laver T, Harrison J, O’Neill PA, et al. Assessing the performance of the Oxford Nanopore Technologies MinION. Biomol Detect Quantif 2015; 3 : 1-8. [Google Scholar]
- Zhao Y, Ashcroft B, Zhang P, et al. Single-molecule spectroscopy of amino acids and peptides by recognition tunnelling. Nat Nanotechnol. 2014; 9 : 466-73. [Google Scholar]
- Wanunu M. Nanopores: a journey towards DNA sequencing. Phys Life Rev 2012; 9 : 125-58. [Google Scholar]
- Feng J, Liu K, Bulushev RD, et al. Identification of single nucleotides in MoS2 nanopores. Nat Nanotechnol 2015; 10 : 1070-6. [Google Scholar]
- Heerema SJ, Dekker C. Graphene nanodevices for DNA sequencing. Nat Nanotechnol 2016; 11 : 127-36. [Google Scholar]
- Schneider GF, Kowalczyk SW, Calado VE, et al. DNA translocation through graphene nanopores. Nano Lett 2010; 10 : 3163-7. [Google Scholar]
- Merchant CA, Healy K, Wanunu M, et al. DNA translocation through graphene nanopores. Nano Lett 2010; 10 : 2915-21. [Google Scholar]
- Hemamouche A, Morin A, Bourhis E, et al. FIB patterning of dielectric, metallized and graphene membranes: A comparative study. Microelectron Eng 2014; 121 : 87-91. [Google Scholar]
- Yusko EC, Johnson JM, Majd S, et al. Controlling protein translocation through nanopores with bio-inspired fluid walls. Nat Nanotechnol 2011; 6 : 253-60. [Google Scholar]
- McNally B, Singer A, Yu Z, et al. Optical recognition of converted DNA nucleotides for single-molecule dna sequencing using nanopore arrays. Nano Lett 2010; 10 : 2237-44. [Google Scholar]
- Levene MJ, Korlach J, Turner SW, et al. zero-mode waveguides for singlrmolecule analysis at high concentrations. Science 2003; 299 : 682-6. [Google Scholar]
- Gilboa T, Meller A. Optical sensing and analyte manipulation in solid-state nanopores. Analyst 2015; 140 : 4733-47. [Google Scholar]
- Rosenstein JK, Wanunu M, Merchant CA, et al. Integrated nanopore sensing platform with sub-microsecond temporal resolution. 2012; 9 : 487-92. [Google Scholar]
- Auger T, Mathé J, Viasnoff V, et al. Zero-mode waveguide detection of flowdriven DNA translocation through nanopores. Phys Rev Lett 2014; 113 : 28302. [Google Scholar]
- Jordan B. Séquençage d’ADN : l’offensive des nanopores. Med Sci (Paris) 2017; 33 : 801-4. [CrossRef] [EDP Sciences] [PubMed] [Google Scholar]
Liste des figures
|
Figure 1. Séquençage par nanopore biologique. L’ADN à séquencer est capté par une enzyme (hélicase ou ADN polymérase, en bleu) qui extrude l’ADN simple brin à une vitesse réduite. Le nanopore lui-même (en rouge) constitue une ouverture dans la membrane lipidique (en vert) dans laquelle le champ électrique (flèches bleues) force le passage de l’ADN simple brin (en marron). Ce dernier est ralenti au passage par la constriction du nanopore (encart du bas) et permet l’identification des bases individuelles par mesure de l’intensité du courant électrique traversant le pore à un instant donné (encart du haut). Le trait pointillé bleu représente le courant à vide du nanopore. |
| Dans le texte | |
|
Figure 2. Séquençage par nanopore artificiel. L’ADN simple brin (ADNsb en marron) est entraîné par le champ électrique (flèches bleues) à travers un nanopore formé par perçage (faisceau d’électrons issus d’un microscope électronique à transmission [TEM] ou faisceau d’ions [FIB]) dans (A) une membrane ultrafine en nitrure de silicium (SiN), ou (B) un matériau bidimensionnel (disulfure de molybdène [MoS2], graphène, nitrure de bore [BN]). L’identification des bases se fait par la mesure du courant ionique traversant le nanopore. |
| Dans le texte | |
|
Figure 3. Séquençage par nanopore artificiel et détection optique. L’ADN est entraîné par un champ électrique ou un flux hydrodynamique à l’intérieur d’un nanopore artificiel. Lorsque qu’une portion de l’ADN atteint l’extrémité du pore, elle est illuminée par un champ électromagnétique évanescent (en vert) qui induit la fluorescence séquentielle de chacune des paires de bases. Les bases sont alors identifiées par un marqueur fluorescent spécifique. ADNsb : ADN simple brin. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.