")
")
| Issue |
Med Sci (Paris)
Volume 39, Number 8-9, Août–Septembre 2023
|
|
|---|---|---|
| Page(s) | 665 - 667 | |
| Section | Forum | |
| DOI | https://doi.org/10.1051/medsci/2023090 | |
| Published online | 11 September 2023 | |
Tout savoir sur les mutations germinales chez les vertébrés
Chroniques génomiques
Everything about germinal mutations in vertebrates
Biologiste, généticien et immunologiste, Président d’Aprogène (Association pour la promotion de la Génomique), 13007 Marseille, France
* This email address is being protected from spambots. You need JavaScript enabled to view it.
Abstract
Mutations in the germline are an essential component of evolution. Current DNA sequencing systems allow quantification of these mutations through full genome sequencing of father/mother/child trios. A recent paper reports such a large-scale study spanning 68 vertebrate species, and establishes important features of the mutation process.
© 2023 médecine/sciences – Inserm
 Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l'utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l'utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.

Dans une chronique récente [1] (→) nous avions analysé les résultats d’études mesurant le taux de mutations somatiques [2], ces mutations qui apparaissent dans les cellules de l’organisme et sont impliquées notamment dans le vieillissement cellulaire et la cancérogenèse, mais ne sont pas transmises à la descendance. On constatait notamment que des animaux de longévités très différentes ont, vers la fin de leur vie, accumulé un nombre comparable de mutations somatiques (quelques milliers). La présente chronique concerne, elle, les mutations germinales, celles qui apparaissent dans les oocytes et les spermatozoïdes et sont retrouvées dans l’ADN de la descendance. Ces mutations sont très importantes car elles sont responsables de l’évolution des espèces et, parfois, provoquent l’apparition de maladies génétiques. Leur mesure est relativement aisée puisqu’il « suffit » de comparer les séquences intégrales d’ADN de trios (père/mère/enfant) : si ces séquences sont de bonne qualité et que leur analyse informatique est bien conduite, on pourra repérer les mutations apparues de novo, absentes chez les parents et présentes dans leur descendance [3]. Du point de vue expérimental, cette analyse est nettement moins ardue que la détection de mutations somatiques qui est compliquée par l’hétérogénéité des cellules analysées [1, 2]. Pour fixer les idées, disons que chez l’homme, le nombre de mutations germinales est de quelques dizaines par génération. La plupart de ces mutations n’ont aucun effet, mais certaines peuvent, rarement, inactiver un gène ou changer certaines propriétés de la protéine que celui-là code.
(→) Voir la Chronique génomique de B. Jordan, m/s n° 8-9, août-septembre 2022, page 723
Une étude de grande ampleur
L’article qui fait l’objet de cette chronique [4] émane d’un ensemble de laboratoires européens et chinois, notamment BGI (Beijing Genomics Institute) [5] qui a réalisé l’ensemble des séquences à l’aide des machines BGI/MGI1 [5] (→).
→ Voir la Chronique génomique de B. Jordan, m/s n° 5, mai 2023, page 469
Les auteurs ont choisi 68 espèces de vertébrés (de l’alpaca Vicugna pacos à l’orque Orcinus orca en passant par l’homme et la souris) pour lesquels une séquence de référence est disponible, ce qui permet d’assembler les courtes séquences produites par les machines2. Ils ont identifié 151 trios (souvent deux ou trois trios par espèce) pour lesquels les informations (notamment l’âge à la reproduction) et l’ADN étaient disponibles, ont vérifié les parentés par analyse d’ADN, et procédé à un séquençage intégral et soigné de ces 453 (151 × 3) génomes : la redondance moyenne de lecture était de 70 fois, alors qu’on se contente généralement de 30 fois. Les séquences obtenues pour chacun des trios ont ensuite été comparées selon un schéma très sophistiqué [6] pour éliminer autant que possible les artefacts, avec un ensemble de contrôles qui donne confiance dans les résultats.
Quelques résultats
Le résultat le plus frappant de cette comparaison d’un ensemble d’animaux très divers est que le nombre de mutations germinales par génération est du même ordre de grandeur dans tous les cas, alors que les masses des animaux varient d’un facteur de plus d’un million (entre l’orque et le poisson combattant chinois [Betta splendens]) et que leur temps de génération va de six mois (pour le chat et le porc) à une trentaine d’années (pour notre espèce et pour certains oiseaux). La Figure 1 montre l’ensemble des données : le taux de mutation par site et par génération augmente un peu avec l’âge à la reproduction mais reste assez proche de 10– 8 par site par génération, soit environ 60 mutations germinales pour un génome humain diploïde de 6 000 mégabases (6 × 109 bases).
 |
Figure 1. Taux moyen de mutations germinales par génération en fonction de l’âge à la reproduction (valeur similaire à celle du temps de geìneìration). On voit que ce taux augmente (leìgèrement) avec l’âge à la reproduction, mais qu’il reste du même ordre pour toutes les 68 espèces. Le groupe auquel appartient l’espèce est donneì par la couleur du point ; le point « oiseau » le plus à droite correspond à un couple de pingouins empereur (Aptenodytes forsteri) dont les deux partenaires eìtaient âgeìs de 36 ans lors de la reproduction (extrait partiel et modifieì de la figure 1 de [4]). |
On retrouve aussi, dans cette étude très large, un phénomène déjà connu, la prédominance des mutations germinales provenant des mâles [7]. Cela correspond au fait que les spermatozoïdes sont produits tout au long de l’existence, alors que le « stock » d’oocytes est constitué dès la naissance. Les résultats montrent que cet excès est assez variable selon les groupes d’espèces : faible chez les reptiles et les poissons, il est de l’ordre d’un facteur trois chez les primates, et supérieur à six chez certains oiseaux (notamment les passereaux). Reptiles et poissons ont généralement des périodes de reproduction courtes et espacées, avec une production de spermatozoïdes par intermittence et donc un nombre de divisions cellulaires réduit par rapport à une production continue, ce qui a pour conséquence une moindre accumulation de mutations germinales chez les mâles.
Il a été proposé que le taux de mutations germinales dépend de la taille effective de la population3 [8], la sélection étant plus efficace sur de grandes populations et tendant ainsi à réduire ce taux. Cette taille effective peut être estimée à partir du taux de substitutions [9], et la Figure 2 montre effectivement une (faible) corrélation négative entre ce paramètre et le taux de mutations par génération, comme attendu.
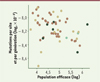 |
Figure 2. Rapport entre le taux de mutation par génération et la taille effective de la population. Les codes de couleur sont identiques à ceux de la Figure 1. La correìlation est relativement marginale (extrait partiel et modifieì de la figure 3 de [4]). |
Si l’on s’intéresse enfin au taux de mutations par année (et non plus par génération), ce paramètre est très différent d’une espèce à l’autre. Il est notamment bien plus élevé chez les espèces domestiquées que chez les espèces sauvages ; les auteurs montrent que cette différence ne provient vraisemblablement pas d’un changement du processus de mutation dû aux conditions de l’élevage, mais de la diminution de l’âge de reproduction et donc du temps de génération : un nombre équivalent de mutations est ainsi concentré sur une durée plus courte.
Les secrets des mutations
Si l’ADN est au centre de la vie, ses mutations sont le matériau de l’évolution, sans laquelle on ne peut rien comprendre à la biologie (« Nothing in biology makes sense except in the light of evolution », comme l’écrivait Theodosius Dobzhansky4 il y a cinquante ans [10]). L’étude globale de ces mutations est longtemps restée hors de portée, et pendant des dizaines d’années, on a dû se limiter à l’analyse de celles qui sont à l’origine de maladies génétiques, ou à l’examen de petites zones de l’ADN présentant un intérêt particulier. Mais aujourd’hui, la rapidité, la précision et le coût abordable du séquençage des génomes entiers permettent d’avoir une vision globale de ces processus de variation. L’étude des génomes anciens permet même de remonter dans le temps et de tester directement des hypothèses en génétique des populations. Le catalogue très large établi par Bergeron et al. [4] n’a pas fini d’être utile, et, au-delà de ses premières conclusions, devrait servir de base à de nombreuses recherches.
Liens d’intérêt
L’auteur déclare n’avoir aucun lien d’intérêt concernant les données publiées dans cet article.
BGI/MGI est actuellement le concurrent le plus sérieux du market leader Illumina [5].
Les séquenceurs BGI/MGI, comme les machines Illumina, fournissent des lectures courtes (100 à 150 nucléotides), contrairement à PacBio et à Oxford Nanopore [5].
Le nombre d’individus participant à la reproduction.
Généticien américain d’origine russe (1900-1975), spécialiste de la génétique des populations.
Références
- Jordan B. Mutations somatiques et longévité. Med Sci (Paris) 2022; 38 : 723–6. [CrossRef] [EDP Sciences] [PubMed] [Google Scholar]
- Cagan A, Baez-Ortega A, Brzozowska N, et al. Somatic mutation rates scale with lifespan across mammals. Nature 2022; 604 : 517–24. [CrossRef] [PubMed] [Google Scholar]
- Tatsumoto S, Go Y, Fukuta K, et al. Direct estimation of de novo mutation rates in a chimpanzee parent offspring trio by ultra-deep whole genome sequencing. Sci Rep 2017 ; 7 : 13561. [CrossRef] [PubMed] [Google Scholar]
- Bergeron LA, Besenbacher S, Zheng J, et al. Evolution of the germline mutation rate across vertebrates. Nature 2023; 615; 285–91. [CrossRef] [PubMed] [Google Scholar]
- Jordan B. Séquençage d’ADN, la fin d’un quasi-monople ? Med Sci (Paris) 2023; 39 : 469–73. [CrossRef] [EDP Sciences] [PubMed] [Google Scholar]
- Bergeron LA, Besenbacher S, Bakker J, et al. The germline mutational process in rhesus macaque and its implications for phylogenetic dating. Gigascience 2021; 10; giab029. [PubMed] [Google Scholar]
- de Manuel M, Wu FL, Przeworski M. A paternal bias in germline mutation is widespread in amniotes and can arise independently of cell division numbers. eLife 2022; 11 : e80008. [CrossRef] [PubMed] [Google Scholar]
- Lynch M. Evolution of the mutation rate. Trends Genet 2010 ; 26 : 345–352. [CrossRef] [PubMed] [Google Scholar]
- Wang J, Santiago E, Caballero A. Prediction and estimation of effective population size. Heredity 2016 ; 117 : 193–206. [CrossRef] [PubMed] [Google Scholar]
- Dobzhansky T. Nothing in Biology Makes Sense Except in the Light of Evolution. American Biology Teacher 1973 ; 35 : 125–129. [CrossRef] [Google Scholar]
Liste des figures
|
Figure 1. Taux moyen de mutations germinales par génération en fonction de l’âge à la reproduction (valeur similaire à celle du temps de geìneìration). On voit que ce taux augmente (leìgèrement) avec l’âge à la reproduction, mais qu’il reste du même ordre pour toutes les 68 espèces. Le groupe auquel appartient l’espèce est donneì par la couleur du point ; le point « oiseau » le plus à droite correspond à un couple de pingouins empereur (Aptenodytes forsteri) dont les deux partenaires eìtaient âgeìs de 36 ans lors de la reproduction (extrait partiel et modifieì de la figure 1 de [4]). |
| Dans le texte | |
|
Figure 2. Rapport entre le taux de mutation par génération et la taille effective de la population. Les codes de couleur sont identiques à ceux de la Figure 1. La correìlation est relativement marginale (extrait partiel et modifieì de la figure 3 de [4]). |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.