")
")
Figure 4.
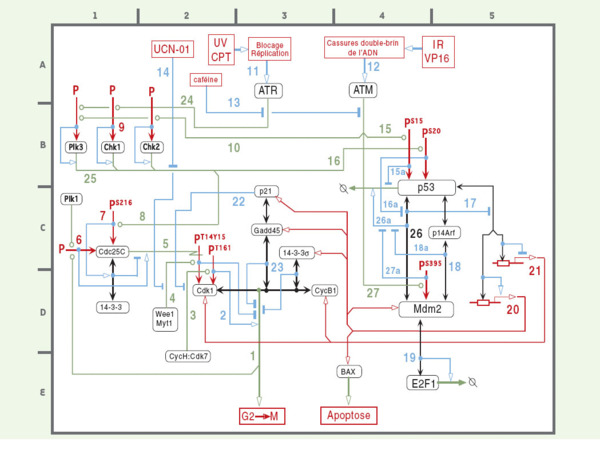
Interactions moléculaires de la transition G2→M. (1)[coordonnées E3] La transition G2→M, qui prépare à la mitose, est principalement contrôlée par le complexe kinase cycline B:Cdk1. (2)[D2] L’activité kinase de Cdk1 est contrôlée par des phosphorylations spécifiques. La phosphorylation de la thréonine 161 est nécessaire pour son activité, alors que la phosphorylation de la thréonine 14 et de la tyrosine 15 sont inhibitrices. (3)[D2] La thréonine 161 de Cdk1 est phosphorylée par la cycline H:Cdk7 (CAK) (voir Figure 3, phosphorylations stimulantes de Cdk2 et Cdk4/6). (4)[D2] La tyrosine kinase nucléaire Wee1 phosphoryle spécifiquement la tyrosine 15 de Cdk1 (et de Cdk2) pendant les phases S et G2, alors que la kinase cytoplasmique Myt1 a une double spécificité pour la thréonine 14 et la tyrosine15 de Cdk1. (5)[C2] Inversement, ces résidus sont déphosphorylés par la phosphatase Cdc25C. (6)[C1] Cdc25C peut être activée par phosphorylation par la Polo kinase Plk1 et par le complexe cycline B:Cdk1, ce qui produit une boucle de contrôle positif pour l’activation de ce complexe. (7)[C1] La phosphorylation de Cdc25C sur la sérine 216 stimule sa liaison à la protéine 14–3–3σ, avec pour résultat une inhibition de sa phosphatase. La liaison de 14–3–3σ à la kinase Weel stimule également l’activité de Wee1 (non montré) [45]. La protéine 14–3–3σ a donc au moins deux mécanismes régulateurs négatifs sur la progression G2→M. (8)[C1-2] La phosphorylation de Cdc25C sur la sérine 216 peut être catalysée par Chk1, Chk2 ou Plk3 (voir annotation 25 ci-dessous). (9)[B1] Chk1 et Chk2 sont activées par phosphorylation. (10)[B2-3] Chk1 semble être principalement phosphorylée par ATR, alors que Chk2 peut être phosphorylée par ATM (ou ATR). (11)[A3] ATR est activée lors de la détection de blocages des fourches de réplication de l’ADN produites par les ultraviolets (UV) ou la camptothécine (CPT). (12)[A4] ATM est activée par des coupures double-brin de l’ADN (DSB) produites par divers agents tels que les radiations ionisantes (IR) ou l’inhibiteur de topo-isomérase II, l’étoposide (VP16). (13)[A-B2-3] La caféine inhibe ATM et ATR. (14)[A2] L’UCN-01 inhibe une variété de protéine kinases, dont Chk1 et Chk2, mais aussi Wee1. Cet effet est indirect. (15)[B4] ATM peut phosphoryler p53 sur sa sérine 15, ce qui stabilise p53 en réduisant sa dégradation. (16)[B3-4] Chk1 et Chk2 phosphorylent p53 sur la sérine 20, ce qui a pour effet de bloquer la liaison de Mdm2. (17)[C5] Mdm2 est un antagoniste de p53, agissant d’une part en l’ubiquitinylant, ce qui déclenche sa dégradation, et d’autre part en bloquant son activité transcriptionnelle. (18)[D5] Mdm2 se fixe à p14Arf, ce qui a pour effet de diminuer la dégradation de p53 liée à la fixation de Mdm2. (19)[E4] Mdm2 se fixe à E2F1 et stimule sa dégradation. Cela bloquerait l’activation des gènes spécifiques de la phase S (voir Figure 3). (20)[D5] La protéine p53 active de nombreux gènes, y compris ceux de Mdm2, p21cip1, Gadd45, 14–3–3 et Bax. (21)[C5] La p53 inhibe d’autre gènes, comme ceux de la cycline B1 et de Cdk1. (22)[C2-3] p21cip1 inhibe plusieurs complexes cycline-kinase (voir Figure 2), comme la cycline H:Cdk7 (CAK). (23)[C-D3] Gadd45 et 14–3–3σ se fixent au complexe cycline B:Cdk1 et bloquent son activité kinase. (24)[A2] Chk1 est activée par les UV et les anomalies de réplication, alors que Chk2 est activée par les radiations ionisantes (IR). (25)[B1] Plk3 est activée par ATM et ATR et, comme Chk1 et Chk2, phosphoryle p53 sur la sérine 20 et Cdc25C sur la sérine 216. Plk3 peut être activée par la doxorubicine, les UV, et le stress oxydatif (H2O2). (26)[C4] Un sillon dans la région N-terminale de Mdm2 lie un segment peptidique dans la région N-terminale de p53, ce qui stimule la dégradation de p53 après ubiquitinylation (26a)[C4]. (27)[D4] ATM inactive l’interaction entre Mdm2 et p53 par phosphorylation de Mdm2.
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.