")
")
| Issue |
Med Sci (Paris)
Volume 35, Number 5, Mai 2019
|
|
|---|---|---|
| Page(s) | 440 - 451 | |
| Section | M/S Revues | |
| DOI | https://doi.org/10.1051/medsci/2019078 | |
| Published online | 22 mai 2019 | |
L’interleukine 33
De la biologie aux implications thérapeutiques
Interleukin-33: from biology to potential treatments
Institut de Pharmacologie et de Biologie Structurale, IPBS, Université de Toulouse, CNRS, UPS, 205, route de Narbonne, 31077 Toulouse, France
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
**
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Résumé
L’interleukine-33 est une cytokine nucléaire de la famille de l’IL-1, exprimée par les cellules endothéliales et épithéliales des tissus en contact avec l’environnement. Elle est libérée lors de dommages tissulaires et joue le rôle d’alarmine en prévenant le système immunitaire d’un danger. Elle est impliquée dans l’immunité innée de type-2 et l’inflammation allergique, mais des études récentes suggèrent qu’elle peut, selon le contexte environnemental, jouer d’autres rôles dans l’homéostasie ou l’immunité antivirale, par exemple. Elle est associée à de nombreuses pathologies, notamment allergiques, inflammatoires ou infectieuses, et pourrait être une cible thérapeutique de choix pour le traitement de l’asthme sévère.
Abstract
Interleukin-33 is a member of the IL-1 cytokine family, expressed in the nucleus of endothelial cells and epithelial cells of barrier tissues. After cellular damage, IL-33 is released in the extracellular space and functions as an alarmin that alerts the immune system. IL-33 plays a critical role in type-2 innate immunity and allergic inflammation, by activating various target cells including mast cells and innate lymphoid cells that secrete high amounts of IL-5 and IL-13, two cytokines involved in allergic reactions. Recent studies suggest that IL-33 can also play other important roles, for example in homeostasis and during viral infection. It is implicated in numerous diseases, including allergic, inflammatory and infectious diseases and it constitutes a promising therapeutic target for treatment of severe asthma.
© 2019 médecine/sciences – Inserm
 Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (http://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l'utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (http://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l'utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.

Vignette [Structure de l’IL-33 (rouge) en complexe avec ST2L (bleu)]. Photo © Protein Data Bank ID:4KC3.
Découverte et structure de l’interleukine 33
L’interleukine 33 (IL-33) (initialement nommée NF-HEV pour nuclear factor of high endothelial venule) a été découverte comme un facteur nucléaire exprimé dans les cellules endothéliales des vaisseaux sanguins HEV1 chez l’homme [1]. Quelques années plus tard, elle a été rattachée aux membres de la famille des interleukines 1 (IL-1) en raison d’une homologie structurale de sa partie C-terminale (acides aminés [aa] 112 à 270) [2]. En effet, ce domaine présente une structure tridimentionnelle en 12 brins bêta organisés en trèfle bêta, caractéristique de l’IL-1 [3] et est doté de l’activité cytokine de la protéine [4]. Sa partie amino-terminale (aa 1 à 78) est nécessaire et suffisante pour amener l’IL-33 dans le noyau et permettre son attachement à la chromatine [5]. Elle comprend une séquence présumée de localisation nucléaire (ou NLS, correspondant aux aa 61 à 78) et un motif de liaison à la chromatine (CBM, aa 40 à 58). Ce motif (MXLRSG) a été identifié comme interagissant avec la poche acide formée par le dimère d’histones H2A-H2B, à la surface du nucléosome [6]. Les domaines N- et C-terminaux de la protéine IL-33 sont conservés au cours de l’évolution (respectivement 59 et 57 % d’identité entre l’homme et la souris), suggérant des fonctions importantes de ces deux domaines. Le domaine central de la protéine (aa 79 à 111 chez l’homme) est une plateforme de clivage et d’activation pour un grand nombre de protéases, endogènes ou exogènes, libérées au cours d’une inflammation allergique ou d’une infection [7-9].
L’axe IL-33/ST2L : de l’expression à l’activation des cellules cibles
Expression de l’IL-33
La séquence de l’IL-33 a été établie dans plus de 110 espèces appartenant toutes à la classe des mammifères. Chez l’homme, l’IL-33 est produite de façon constitutive en condition basale, essentiellement au niveau des vaisseaux sanguins et dans les tissus muqueux exposés à l’environnement extérieur qui constituent la première ligne de défense contre les pathogènes [10]. On la retrouve notamment dans le noyau des cellules endothéliales tout le long de l’arbre vasculaire, des cellules épithéliales de tissus en contact avec l’extérieur (glande salivaire, poumons, estomac, intestin, vagin, etc.), mais aussi dans des fibroblastes (fibroblastic reticular cells, ou FRC) des organes lymphoïdes secondaires [10] et des cellules mésenchymateuses. L’expression de l’IL-33 humaine peut également être induite dans un contexte pathologique qu’il soit d’origine inflammatoire, fibrotique ou infectieux, non seulement dans ces mêmes cellules mais aussi dans d’autres types cellulaires comme les kératinocytes de la peau ou des myofibroblastes au niveau du foie, du pancréas ou des reins [11, 12]. Une expression similaire à celle trouvée chez l’homme a été observée chez la souris à l’exception, notamment, de l’endothélium où seule une expression inductible (lors d’une réaction inflammatoire par exemple) a pu être mise en évidence [13]. D’autres différences d’expression entre l’homme et la souris ont été décrites, par exemple au niveau des cellules épithéliales des voies respiratoires puisque la forme murine est exprimée dans les cellules alvéolaires de type II tandis que chez l’homme, elle est retrouvée dans les cellules épithéliales basales bronchiques [14].
Libération de l’IL-33
L’IL-33 ne possède pas de peptide signal. Elle n’est donc pas sécrétée dans le milieu extracellulaire par la voie de sécrétion classique qui emprunte la route du réticulum endoplasmique et de l’appareil de Golgi. Contrairement à d’autres membres de la famille de l’IL-1, l’IL-33 ne nécessite pas de clivage par la caspase-1 pour être libérée sous forme active dans le milieu extracellulaire [15]. Il est vraisemblable qu’elle soit rapidement libérée de façon passive par les cellules nécrotiques, endommagées à la suite d’un stress tissulaire ou cellulaire, au cours d’une inflammation, d’une infection ou après une lésion. Cependant, il n’est pas exclu que des mécanismes de sécrétion active permettent la libération de l’IL-33 dans le milieu extracellulaire sans qu’il y ait véritablement de mort cellulaire. Différents mécanismes de sécrétion active ont ainsi été proposés, comme le stress oxydatif, l’activation cellulaire par l’ATP, l’acide urique ou les récepteurs PAR (récepteurs activés par les protéases) impliquant notamment une élévation du calcium intracellulaire [16].
Le récepteur de l’IL-33, ST2L, et les voies de signalisation induites dans les cellules cibles
Le récepteur ST2L (suppressor of tumorigenicity 2) est codé par le gène IL1RL1 (interleukin-1 receptor-like 1) et est présent à la surface de nombreuses cellules du système immunitaire, son expression pouvant être constitutive ou induite par l’environnement [12]. Les cellules cibles majeures de l’IL-33 sont les cellules immunitaires innées qui résident dans les tissus et expriment le récepteur ST2L de façon constitutive comme les cellules lymphoïdes innées de type 2 (ILC2), les mastocytes, les basophiles, les macrophages et les lymphocytes T régulateurs (Treg) [16-19] (→) qui sont capables de répondre très rapidement à la libération d’IL-33 dans les tissus. Les voies de signalisation activées par l’IL-33 (Figure 1) dans les cellules cibles sont très similaires à celles activées par l’IL-1 et l’IL-18 [4]. L’expression différentielle des récepteurs ST2L, IL-1R et IL-18R (les récepteurs des IL-1 et IL-18) sur les cellules cibles est ainsi critique pour expliquer les effets biologiques uniques de chaque cytokine.
(→) Voir la Synthèse de A. Crinier et al., m/s n° 5, mai 2017, page 534
 |
Figure 1. Voies de signalisation de l’IL-33. Le récepteur ST2L (suppressor of tumorigenicity 2) comprend une partie extracellulaire composée de trois domaines Ig-like, un domaine transmembranaire et un domaine cytoplasmique TIR (Toll-likeinterleukin 1 receptor). La liaison de l’IL-33 à son récepteur ST2L entraîne un changement conformationnel qui permet la formation d’un complexe trimérique de haute affinité avec le corécepteur IL1RAcP (IL-1 receptor accessory protein), également corécepteur de l’IL-1 (interleukine 1) [4]. Comme pour l’IL-1, le rapprochement des deux domaines TIR entraîne le recrutement de la protéine adaptatrice MyD88 (myeloid differentiation primary response protein 88) qui initie la cascade de signalisation impliquant les molécules adaptatrices IRAK1 (IL-1b receptor-associated kinase 1), IRAK4 (IL-1b receptor-associated kinase 4), et TRAF6 (TNF receptor associated factor 6) [2]. Les voies IKK (IκB kinase)/NF-κB (nuclear factor-kappa B) et MAPK (mitogen-activated protein kinase) (p38, JNK [c-Jun N-terminal kinase] ou ERK [extracellular signal-regulated kinase]) sont alors activées, permettant notamment la translocation des facteurs de transcription AP-1 (activator protein 1) et NF-κB et la transcription de gènes cibles. |
Fonctions biologiques de l’IL-33 et pathologies associées
L’expression constitutive de l’IL-33 au niveau de l’endothélium et des barrières épithéliales en fait une cytokine de choix pour initier rapidement les réponses immunes à la suite de dommages causés par divers agents pathogènes (virus, parasites) ou environnementaux (allergènes). L’IL-33 fonctionne comme un signal d’alarme (ou alarmine), qui sent le dommage tissulaire et alerte les cellules du système immunitaire.
Rôle majeur de l’IL-33 dans les réponses immunes de type 2 et les maladies allergiques
De nombreuses études ont permis de montrer que l’administration exogène d’IL-33 à des souris, par exemple par voie intra-nasale ou péritonéale, entraîne l’activation et l’expansion de cellules immunitaires innées cibles de l’IL-33, notamment les ILC2, les mastocytes et les basophiles [19]. Cette activation conduit à la production massive de cytokines de type 2 (IL-4, IL-5 et IL-13), de chimiokines (éotaxine), et de médiateurs pro-inflammatoires (IL-6, protéases, histamine, eicosanoïdes, etc.). Une immunité de type 2 est alors mise en place, entraînant une éosinophilie, l’hyperplasie des cellules caliciformes, la production de mucus, la contraction des muscles lisses et la commutation isotypique2 vers la synthèse d’immunoglobulines d’isotype E (IgE) [16, 17] (Figure 2). Ces réponses sont également observées chez des souris déficientes en lymphocytes B et T (les souris Rag2-/- [recombination activating gene 2]) suggérant que les cellules innées sont des cibles majeures de l’IL-33. D’autres cellules immunitaires innées (éosinophiles, macrophages alvéolaires de phénotype M2, cellules dendritiques, iNKT [invariant natural killer T]) ou adaptatives (lymphocytes T CD4+), recrutées sur le site inflammatoire et exprimant le récepteur ST2L de façon constitutive ou inductible, sont également activées et/ou polarisées par l’IL-33 et participent au processus inflammatoire.
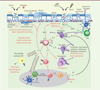 |
Figure 2. Rôles de l’IL- 33 dans l’initiation des réponses immunes de type 2, au niveau des poumons. Suite à divers stimulus qui endommagent la barrière épithéliale du poumon (allergène, lésion, parasite, stress, etc.), l’IL-33 est libérée des cellules épithéliales. Très rapidement, elle active et induit la prolifération des cellules résidentes ILC2 (type 2 innate lymphoid cells), mastocytes et basophiles. Ces cellules sécrètent des cytokines Th2 (IL-4, IL-5, IL-13, IL-9) et divers médiateurs inflammatoires qui permettent la mise en place de la réponse innée de type 2. L’IL-13 favorise la migration des cellules dendritiques (DC) dans les noeuds lymphatiques qui vont permettre la différenciation des lymphocytes T naïfs (T Naïfs) en lymphocyte T helper 2 (Th2). L’IL-13 participe au recrutement, à l’activation et à la polarisation des macrophages alvéolaires M2. L’IL-5 est une cytokine clé pour le recrutement, l’activation et la survie des éosinophiles. L’ensemble de ces signaux conduit à l’inflammation des voies respiratoires, avec l’hyperplasie des cellules caliciformes, la production de mucus (qui permet l’expulsion des vers), l’hyperréactivité pulmonaire, l’éosinophilie et la production d’IgE (immunoglobuline E). Certains des signaux émis (amphiréguline ou AREG) permettent de réparer les lésions et participent au remodelage de l’épithélium. B : lymphocyte B ; Treg : lymphocyte T régulateur ; IL : interleukine. |
Ainsi, l’IL-33 joue un rôle prépondérant dans l’initiation et le développement des réponses immunes de type 2, qui s’avèrent primordiales lors de l’infection de l’hôte par des helminthes, par exemple. En effet, l’administration de l’IL-33 à des souris infestées par ces vers parasites permet de les expulser [20], notamment en induisant les cytokines de type 2, IL-4 et IL-13, et des souris déficientes en IL-33 présentent un défaut d’élimination de ces vers. Chez l’homme, l’expression de l’IL-33 est augmentée dans le sérum de patients infestés par Schistosoma japonica3, et est corrélée à une induction des cytokines de type 2. L’IL-33 exerce ainsi un effet protecteur contre les infestations4 par de nombreux vers parasites. En revanche, cette immunité de type 2, induite par l’IL-33, peut contribuer au développement de diverses maladies allergiques telle que l’asthme, la dermatite atopique, la rhinite allergique ou la rhinosinusite chronique [12]. De nombreuses études réalisées dans divers modèles de rhinite ou d’asthme allergique chez la souris ont ainsi montré que la voie IL-33/ST2L/ILC2 était requise pour l’inflammation allergique des voies aériennes [21-25]. De même, dans un modèle expérimental de dermatite atopique, le développement de la maladie est inhibé chez les souris invalidées pour le gène Il-33 ou Il1rl1. La surexpression de l’IL-33 dans la peau entraîne également des symptômes caractéristiques de la dermatite atopique [26], suggérant un rôle important de la cytokine dans cette pathologie. Ainsi, l’IL-33, en initiant et amplifiant les réponses immunes de type 2, apparaît être un acteur central dans la marche atopique, depuis la dermatite atopique jusqu’à la rhinite allergique et l’asthme [27]. Chez l’homme, l’IL-33 semble impliquée dans ces mêmes pathologies allergiques. En effet, les niveaux d’expression d’IL-33, d’IL-5, d’IL-13 ainsi que le nombre d’éosinophiles et d’ILC2 augmentent chez les patients atteints de dermatite atopique, de rhinite allergique ou d’asthme, et sont corrélés à la sévérité de la maladie [28]. Il a été par ailleurs proposé que l’IL-33 soit un médiateur du remodelage des voies respiratoires chez les enfants atteints d’asthme sévère [29].
Les autres rôles de l’IL-33 (Figure 3)
Dans les réponses immunes de type 1
 |
Figure 3. Les multiples rôles de l’IL-33 dans le système immunitaire. L’IL-33 est libérée des cellules productrices, majoritairement des cellules épithéliales et endothéliales, dans le milieu extracellulaire par divers stimuli (parasite, virus, allergène, lésion, etc.). Elle active et induit la prolifération ou permet la polarisation de nombreuses cellules cibles qui expriment le récepteur ST2L dans un contexte donné. L’ensemble des médiateurs sécrétés (cytokines, chimiokines ou autre) et les différentes interactions cellulaires qui sont alors établies vont permettre d’orienter la réponse immune. Ainsi, selon le tissu et l’environnement, l’IL-33 peut jouer des rôles très différents, et être impliquée dans des réponses pro- ou anti-inflammatoires, dans l’immunité antiparasitaire, antivirale, dans les processus de réparation tissulaire, etc. Des études récentes ont montré que l’IL-33 participait également à la genèse et/ou au maintien de l’homéostasie de différents tissus [38-40]. |
Un nombre croissant d’études montre que l’IL-33 présente des propriétés immunologiques différentes en fonction du microenvironnement qui module la distribution du récepteur ST2L et le type de cellules activées. Elle peut notamment promouvoir des réponses immunitaires de type 1, par exemple dans un contexte infectieux et/ou d’inflammation chronique. Ainsi, l’IL-33, en combinaison avec l’IL-12 produite en réponse à une infection virale, contribue à l’expansion et à l’activation des cellules NK et iNKT, en induisant la production d’interféron-g (IFN-g) [30]. De même, l’IL-33 et l’IL-12 coopèrent pour permettre l’augmentation transitoire de l’expression du récepteur ST2L sur les lymphocytes T cytotoxiques (CTL) CD8+, et stimulent l’expansion et la fonction effectrice des CTL activés ainsi que la sécrétion des cytokines antivirales, telles que l’IL-10 ou l’IFN-g [31]. Ainsi, l’IL-33 joue un rôle important dans l’immunité antivirale protectrice. Dans un contexte de bronchopneumopathie chronique obstructive (BPCO), l’IL-33 entraîne la sécrétion de cytokines pro-inflammatoires comme le TNF-a (tumor necrosis factor), l’IL-12 et l’IFN-g, par les macrophages et les cellules NK, augmentant ainsi les réponses pro-inflammatoires de type 1 et exacerbant la maladie inflammatoire chronique [32].
Dans la résolution de l’inflammation, la réparation tissulaire et l’homéostasie
Même si l’IL-33 est impliquée dans les processus inflammatoires, elle contribue également à la résolution de l’inflammation et permet le retour à l’homéostasie en participant au processus de réparation tissulaire. Par exemple, il a été démontré que la signalisation induite par la liaison de l’IL-33 à son récepteur ST2L était requise pour restaurer l’intégrité de l’épithélium des voies respiratoires, après une infection des poumons par le virus de l’influenza ou après une infestation par le nématode Nippostrongylus brasiliensis 5 [11]. Cette réparation est due en partie à l’amphiréguline, un facteur de croissance sécrété notamment par les ILC2, les mastocytes et les lymphocytes T régulateurs (Treg) et qui se lie au récepteur EGFR (epidermal growth factor receptor) [33, 34]. D’autres évidences suggèrent que l’IL-33 est également un important régulateur de la réparation tissulaire dans la peau, les muscles, le foie et l’intestin, en permettant l’expansion et la fonction effectrice des lymphocytes Treg, directement ou indirectement via l’activation des ILC2 et la polarisation des macrophages6. Cependant, lorsque ce processus de réparation est mal contrôlé, il peut contribuer au développement de fibrose dans différents organes [35].
L’IL-33 semble jouer un rôle important dans l’homéostasie du poumon puisque qu’elle permet l’expansion et l’activation des ILC2 dans les poumons des nouveau-nés [36, 37]. Les réponses immunes de type 2 qui en résultent sont préservées tout au long de la vie et, à l’homéostasie, elles maintiennent les macrophages alvéolaires dans un phénotype de type M2 [37], induisent un état d’activation basal des ILC2 [38] et modulent le transcriptome des basophiles résidant dans les poumons [39]. L’IL-33 est également requise pour le maintien de l’homéostasie dans le tissu adipeux, en facilitant la différenciation et le maintien des lymphocytes Treg Foxp3+ ST2+ et des ILC2 dans le tissu adipeux viscéral. Cela a pour conséquence de limiter l’inflammation associée à l’obésité et de préserver la sensibilité à l’insuline ainsi que la tolérance au glucose [40].
L’IL-33 et les pathologies
Au vu du large spectre d’expression de l’IL-33, de ses cellules cibles et de ses fonctions, il n’est pas étonnant que l’IL-33 soit associée à de nombreuses pathologies, non seulement les maladies allergiques comme évoqué plus haut, mais également les pathologies inflammatoires chroniques et fibrotiques touchant l’intestin (rectocolite hémorragique ou colite ulcéreuse) ou le poumon (BPCO), les maladies infectieuses (sepsis, infection par des vers, virus ou bactéries), et les maladies cardiovasculaires (athérosclérose), rénales et métaboliques [12]. L’IL-33 joue tantôt un rôle protecteur, tantôt un rôle néfaste dans ces pathologies (Tableau I).
Exemples de l’implication de l’axe IL-33/ST2 dans les maladies [12]. IL-33 : interleukine-33 ; ILC2 : cellules lymphoïdes innées de type 2 ; ST2 : suppression of tumorigenicity 2 ; sST2 : ST2 soluble ; NK : natural killer ; LT CD8+ : lymphocyte T CD8+ ; Th1 : lymphocyte T helper 1 ; DC : cellules dendritiques ; Th2 : lymphocyte T helper 2, Il1rl1 : Interleukin 1 receptor-like 1 ; IL-13 : interleukine 13 ; BPCO : bronchopneumopathie chronique obstructive ; DSS : dextran sodium sulfate ; TNBS : trinitrobenzene sulfonic acid ; Treg : lymphocytes T régulateurs ; RGCH : réaction du greffon contre l’hôte.
Régulation de l’IL-33
La protéine IL-33 doit être finement régulée afin qu’elle puisse exercer pleinement ses fonctions en cas de danger, sans devenir néfaste en participant au développement de diverses pathologies aiguës ou chroniques. Plusieurs mécanismes moléculaires sont impliqués dans la régulation de son activité.
Séquestration de l’IL-33 dans le noyau
Dans un modèle de souris knock-in, dans lequel le domaine amino-terminal de l’IL-33, comprenant le CBM, a été éliminé, l’IL-33 n’est plus séquestrée dans le noyau de la cellule. Elle est alors libérée de façon constitutive dans le milieu extra-cellulaire et sécrétée dans le sérum, ce qui induit une inflammation massive au niveau de nombreux organes [41]. Cette inflammation est dépendante du récepteur ST2L et entraîne la mort des souris à l’âge de trois mois. Plusieurs études de transcriptomique et protéomique ont démontré, tant in vitro qu’in vivo, que l’IL-33 localisée dans le noyau de cellules endothéliales ou épithéliales n’avait pas d’effet sur la transcription des gènes. En revanche, l’IL-33 exogène joue son rôle de cytokine [42]. Ainsi, l’IL-33 n’aurait pas de rôle actif au sein du noyau, mais y serait retenue pour limiter sa fonction de cytokine inflammatoire extra-cellulaire. Cette localisation dans le noyau a été conservée au cours de l’évolution probablement pour maintenir l’homéostasie tissulaire et protéger l’organisme d’une inflammation létale non contrôlée.
Séquestration de l’IL-33 par le récepteur soluble sST2
Deux isoformes majeures du récepteur ST2 ont été décrites : la forme transmembranaire (ST2L) et une forme soluble (sST2) qui ne possède que le domaine extracellulaire. La forme soluble sST2 est sécrétée par de nombreux types cellulaires, notamment les mastocytes et les lymphocytes T de type Th2. Elle agit comme un récepteur leurre capable de neutraliser l’IL-33 libre dans les fluides biologiques [21], empêchant ainsi les effets systémiques de la cytokine dans le sang. De hauts niveaux de sST2 sont retrouvés dans le sérum des patients atteints de maladies inflammatoires. Ils sont généralement corrélés à la sévérité de la maladie.
Inactivation de l’IL-33 par oxydation
L’activité biologique de l’IL-33 est régulée par oxydation [43]. Elle est libérée dans le milieu extracellulaire sous sa forme réduite et est rapidement (en moins de 2 heures) oxydée au niveau de quatre cystéines situées dans son domaine C-terminal (Cys208, Cys227, Cys232 et Cys259). Cette oxydation induit la formation de deux ponts disulfures et un changement conformationnel de la protéine qui entraîne son inactivation. La mutation de ces quatre cystéines a pour effet de prolonger l’activité de l’IL-33 dans le poumon. Les premières heures qui suivent sa libération dans le milieu extracellulaire sont donc critiques pour l’activation des cellules cibles.
Régulation de l’IL-33 par les protéases
L’IL-33 est un substrat de nombreuses protéases qui régulent son activité, que ce soit en l’inhibant ou en l’activant. Plusieurs études ont démontré que les caspases apoptotiques 3 et 7 clivent l’IL-33 dans son domaine cytokine et l’inactivent, probablement afin qu’elle ne provoque pas de réaction inflammatoire inopportune au cours de la mort cellulaire programmée [15]. D’autres travaux ont montré que les protéases endogènes produites par les mastocytes (chymase, tryptase) et les neutrophiles (cathepsine G, élastase et protéinase 3), clivent l’IL-33 dans son domaine central [7, 8] générant ainsi des formes matures biologiquement « superactives ». Une étude récente a démontré que l’IL-33 est un véritable détecteur (sensor) des activités protéolytiques associées aux allergènes environnementaux, tels que les pollens, les champignons, les acariens, etc. [9]. En effet, elle est clivée dans son domaine central par de nombreuses protéases exogènes provenant d’allergènes. Les fragments C-terminaux de la protéine ainsi générés présentent une activité au moins 30 fois supérieure à celle de la forme complète, conduisant à l’initiation de l’inflammation allergique. Ces clivages pourraient avoir un rôle déterminant in vivo comme facteur déclenchant de réactions allergiques, en particulier lorsque l’IL-33 est présente à de faibles concentrations.
Implications thérapeutiques : de nouveaux traitements en perspective pour l’asthme sévère et les maladies allergiques
De nombreuses études génétiques ont montré que l’IL-33 joue un rôle majeur dans l’asthme chez l’homme. En effet, les gènes IL33 et IL1RL1 ont été identifiés, de façon récurrente, comme des gènes de susceptibilité majeurs à l’asthme dans des études de très grande envergure (genome wide association studies, GWAS) impliquant plus de 10 000 patients d’origines ethniques différentes et présentant différentes formes d’asthme [16, 44, 45]. La mutation d’un seul allèle, qui entraîne une perte de fonction de l’IL-33, réduit ainsi le nombre d’éosinophiles dans le sang et protège de l’asthme [46]. D’autres gènes de susceptibilité à l’asthme jouant des rôles importants dans la différenciation des ILC2 (RORA [RAR-related orphan receptor A]), leur prolifération (IL2RB [interleukin 2 receptor subunit beta]), leur activation (TSLP [thymic stromal lymphopoietin], IL2RB) et leur fonction (IL13), ont été identifiés dans diverses études génétiques, indiquant que l’axe IL-33/ST2/ILC2 joue un rôle crucial dans l’asthme [45]. De plus, le fait que l’IL-33 soit libérée très rapidement après exposition à un allergène [9], qu’elle soit impliquée en amont de la cascade des réactions allergiques, et qu’elle soit active dans différentes étapes de la maladie atopique, suggère qu’elle est une cible thérapeutique de choix pour le traitement de l’asthme et, de façon plus générale, des maladies allergiques. D’ailleurs, l’inhibition de la voie IL-33/ST2 permet d’améliorer les symptômes de la maladie allergique dans des modèles murins de rhinite allergique [47], d’asthme allergique, d’allergie alimentaire (phase de sensibilisation) et de dermatite atopique. Chez l’homme, de nouveaux traitements des maladies allergiques fondés sur l’utilisation d’anticorps monoclonaux ciblant la voie IL-33/ST2 sont en phases d’essais cliniques et semblent très prometteurs (Tableau II).
Anticorps monoclonaux thérapeutiques dans les maladies allergiques. Moduler les acteurs majeurs des réponses de type 2 représente une stratégie prometteuse pour traiter les maladies allergiques. Des thérapies à base d’anticorps visant à bloquer les IgE ou les cytokines de type 2 ont déjà été utilisées durant cette dernière décennie pour lutter contre l’asthme sévère et la dermatite atopique. Plusieurs grandes sociétés pharmaceutiques développent maintenant des anticorps monoclonaux ciblant directement la voie IL-33/ST2 dans l’espoir d’améliorer les traitements existants et d’étendre le spectre des maladies concernées. Les premiers résultats avec ces anticorps anti-IL-33/ST2, actuellement en essai clinique de phase I ou II, sont très encourageants. FDA : food and drug administration ; AMM : autorisation de mise sur le marché.
Conclusion et perspectives
De plus en plus de travaux montrent que l’IL-33, au-delà de sa fonction d’alarmine, se comporte comme une cytokine pléiotrope, n’agissant pas de façon isolée. En effet, d’autres facteurs (cytokines, médiateurs lipidiques, neuropeptides, etc.) peuvent amplifier, diminuer, complexifier ou détourner son action. Étudier la coopération de l’IL-33 avec d’autres régulateurs en homéostasie ou dans des contextes pathologiques devrait permettre d’obtenir une vue plus complète et complexe de ses fonctions. Même si de grands pas ont été réalisés ces dernières années, notamment la découverte des cellules cibles ILC2, certaines interrogations subsistent, en particulier : quels sont les facteurs qui régulent l’expression de l’IL-33 ? Comment l’IL-33 est-elle libérée dans le milieu extracellulaire ? Pourquoi ne sommes-nous pas tous allergiques ? Comment réconcilier le rôle de l’IL-33 tantôt néfaste, tantôt protecteur, dans différentes pathologies qui, parfois, coexistent au sein d’un individu, comme dans l’asthme associé à l’obésité par exemple ?
Tant de questions auxquelles nous n’avons pas encore de réponse. Cependant, l’intérêt grandissant des biologistes et des entreprises pharmaceutiques pour l’IL-33, soutenu par les avancées technologiques, notamment le séquençage à haut débit sur cellule unique réalisé dans différents contextes, devraient permettre de répondre rapidement à de telles questions.
Liens d’intérêt
Les auteurs déclarent n’avoir aucun lien d’intérêt concernant les données publiées dans cet article.
Remerciements
Ce travail est soutenu par l’Agence nationale de la recherche (ANR-12-BSV3-0005-01 et ANR-16-CE15-0009-01 à J.-P.G.). P. Schmitt est financée par des bourses du ministère de l’Enseignement supérieur, de la Recherche et de l’Innovation, et de la Fondation ARC pour la Recherche sur le Cancer (ARC DOC20180507586).
Ou veinules postcapillaires à endothélium épais. Ce sont des vaisseaux permettant aux lymphocytes d’entrer dans les ganglions lymphatiques et dans certains autres organes lymphoïdes secondaires.
Processus qui, lors de la maturation d’un lymphocyte B, permet de modifier l’isotype des immunoglobulines produites.
C’est l’espèce d’helminthe la plus pathogène pour l’homme. Elle est responsable de la bilharziose artério-veineuse.
On parle d’infestation et non d’infection lorsqu’il s’agit de parasites.
N. brasiliensis infeste son hôte en traversant la peau. Il migre vers les poumons où la larve mature, puis vers l’intestin où la forme adulte du ver réside.
Processus par lequel les macrophages adoptent différents programmes fonctionnels en réponse à des signaux du microenvironnement ; on distingue la polarisation classique de type M1 qui est associée à la production de cytokines pro-inflammatoires et la polarisation alternative de type M2 qui participe à la régulation de l’inflammation et au phénomène de réparation tissulaire.
Références
- Baekkevold ES, Roussigne M, Yamanaka T, et al. Molecular characterization of NF-HEV, a nuclear factor preferentially expressed in human high endothelial venules. Am J Pathol. 2003 ; 163 : 69–79. [CrossRef] [PubMed] [Google Scholar]
- Schmitz J, Owyang A, Oldham E, et al. IL-33, an interleukin-1-like cytokine that signals via the IL-1 receptor-related protein ST2 and induces T helper type 2-associated cytokines. Immunity. 2005 ; 23 : 479–490. [CrossRef] [PubMed] [Google Scholar]
- Lingel A, Weiss TM, Niebuhr M, et al. Structure of IL-33 and its interaction with the ST2 and IL-1RAcP receptors: insight into heterotrimeric IL-1 signaling complexes. Structure. 2009 ; 17 : 1398–1410. [CrossRef] [PubMed] [Google Scholar]
- Dinarello CA. Overview of the IL-1 family in innate inflammation and acquired immunity. Immunol Rev. 2018 ; 281 : 8–27. [CrossRef] [PubMed] [Google Scholar]
- Carriere V, Roussel L, Ortega N, et al. IL-33, the IL-1-like cytokine ligand for ST2 receptor, is a chromatin-associated nuclear factor in vivo. Proc Natl Acad Sci USA. 2007 ; 104 : 282–287. [CrossRef] [Google Scholar]
- Roussel L, Erard M, Cayrol C, Girard JP. Molecular mimicry between IL-33 and KSHV for attachment to chromatin through the H2A–H2B acidic pocket. EMBO Rep. 2008 ; 9 : 1006–1012. [Google Scholar]
- Lefrancais E, Roga S, Gautier V, et al. IL-33 is processed into mature bioactive forms by neutrophil elastase and cathepsin G. Proc Natl Acad Sci USA. 2012 ; 109 : 1673–1678. [CrossRef] [Google Scholar]
- Lefrancais E, Duval A, Mirey E, et al. Central domain of IL-33 is cleaved by mast cell proteases for potent activation of group-2 innate lymphoid cells. Proc Natl Acad Sci USA. 2014 ; 111 : 15502–15507. [CrossRef] [Google Scholar]
- Cayrol C, Duval A, Schmitt P, et al. Environmental allergens induce allergic inflammation through proteolytic maturation of IL-33. Nat Immunol. 2018 ; 19 : 375–385. [CrossRef] [PubMed] [Google Scholar]
- Moussion C, Ortega N, Girard JP. The IL-1-like cytokine IL-33 is constitutively expressed in the nucleus of endothelial cells and epithelial cells in vivo: a novel ‘alarmin’?. PloS One. 2008 ; 3 : e3331. [CrossRef] [PubMed] [Google Scholar]
- Molofsky AB, Savage AK, Locksley RM. Interleukin-33 in tissue homeostasis, injury, and inflammation. Immunity. 2015 ; 42 : 1005–1019. [CrossRef] [PubMed] [Google Scholar]
- Liew FY, Girard JP, Turnquist HR. Interleukin-33 in health and disease. Nat Rev Immunol. 2016 ; 16 : 676–689. [CrossRef] [PubMed] [Google Scholar]
- Pichery M, Mirey E, Mercier P, et al. Endogenous IL-33 is highly expressed in mouse epithelial barrier tissues, lymphoid organs, brain, embryos, and inflamed tissues : in situ analysis using a novel Il-33-LacZ gene trap reporter strain. J Immunol. 2012 ; 188 : 3488–3495. [CrossRef] [PubMed] [Google Scholar]
- Byers DE, Alexander-Brett J, Patel AC, et al. Long-term IL-33-producing epithelial progenitor cells in chronic obstructive lung disease. J Clin Invest. 2013 ; 123 : 3967–3982. [CrossRef] [PubMed] [Google Scholar]
- Cayrol C, Girard JP. The IL-1-like cytokine IL-33 is inactivated after maturation by caspase-1. Proc Natl Acad Sci USA. 2009 ; 106 : 9021–9026. [CrossRef] [Google Scholar]
- Drake LY, Kita H. IL-33: biological properties, functions, and roles in airway disease. Immunol Rev. 2017 ; 278 : 173–184. [CrossRef] [PubMed] [Google Scholar]
- Smith DE. IL-33: a tissue derived cytokine pathway involved in allergic inflammation and asthma. Clin Exp Allerg. 2010 ; 40 : 200–208. [CrossRef] [Google Scholar]
- Crinier A, Viant C, Girard-Madoux M, Vivier E. Les cellules lymphoïdes innées. Med Sci (Paris). 2017 ; 33 : 534–542. [CrossRef] [EDP Sciences] [PubMed] [Google Scholar]
- Cayrol C, Girard JP. Interleukin-33 (IL-33): a nuclear cytokine from the IL-1 family. Immunol Rev. 2018 ; 281 : 154–168. [CrossRef] [PubMed] [Google Scholar]
- Maizels RM, McSorley HJ. Regulation of the host immune system by helminth parasites. J Allergy Clin Immunol. 2016 ; 138 : 666–675. [CrossRef] [PubMed] [Google Scholar]
- Hayakawa H, Hayakawa M, Kume A, Tominaga S. Soluble ST2 blocks interleukin-33 signaling in allergic airway inflammation. J Biol Chem. 2007 ; 282 : 26369–26380. [PubMed] [Google Scholar]
- Oboki K, Ohno T, Kajiwara N, et al. IL-33 is a crucial amplifier of innate rather than acquired immunity. Proc Natl Acad Sci USA. 2010 ; 107 : 18581–18586. [CrossRef] [Google Scholar]
- Haenuki Y, Matsushita K, Futatsugi-Yumikura S, et al. A critical role of IL-33 in experimental allergic rhinitis. J Allergy Clin Immunol. 2012 ; 130 : 184–94. e11. [CrossRef] [PubMed] [Google Scholar]
- Halim TY, Steer CA, Matha L, et al. Group 2 innate lymphoid cells are critical for the initiation of adaptive T helper 2 cell-mediated allergic lung inflammation. Immunity. 2014 ; 40 : 425–435. [CrossRef] [PubMed] [Google Scholar]
- Lee HY, Rhee CK, Kang JY, et al. Blockade of IL-33/ST2 ameliorates airway inflammation in a murine model of allergic asthma. Exp Lung Res. 2014 ; 40 : 66–76. [CrossRef] [PubMed] [Google Scholar]
- Imai Y, Yasuda K, Sakaguchi Y, et al. Skin-specific expression of IL-33 activates group 2 innate lymphoid cells and elicits atopic dermatitis-like inflammation in mice. Proc Natl Acad Sci USA. 2013 ; 110 : 13921–13926. [CrossRef] [Google Scholar]
- Han H, Roan F, Ziegler SF. The atopic march: current insights into skin barrier dysfunction and epithelial cell-derived cytokines. Immunol Rev. 2017 ; 278 : 116–130. [CrossRef] [PubMed] [Google Scholar]
- Prefontaine D, Nadigel J, Chouiali F, et al. Increased IL-33 expression by epithelial cells in bronchial asthma. J Allergy Clin immunol. 2010 ; 125 : 752–754. [CrossRef] [PubMed] [Google Scholar]
- Castanhinha S, Sherburn R, Walker S, et al. Pediatric severe asthma with fungal sensitization is mediated by steroid-resistant IL-33. J Allergy Clin immunol. 2015 ; 136 : 312–22. e7. [CrossRef] [PubMed] [Google Scholar]
- Bourgeois E, Van LP, Samson M, et al. The pro-Th2 cytokine IL-33 directly interacts with invariant NKT and NK cells to induce IFN-gamma production. Eur J Immunol Immunol. 2009 ; 39 : 1046–55. [CrossRef] [Google Scholar]
- Bonilla WV, Frohlich A, Senn K, et al. The alarmin interleukin-33 drives protective antiviral CD8+ T cell responses. Science. 2012 ; 335 : 984–989. [Google Scholar]
- Silver JS, Kearley J, Copenhaver AM, et al. Inflammatory triggers associated with exacerbations of COPD orchestrate plasticity of group 2 innate lymphoid cells in the lungs. Nat Immunol. 2016 ; 17 : 626–635. [CrossRef] [PubMed] [Google Scholar]
- Monticelli LA, Sonnenberg GF, Abt MC, et al. Innate lymphoid cells promote lung-tissue homeostasis after infection with influenza virus. Nat Immunol. 2011 ; 12 : 1045–1054. [CrossRef] [PubMed] [Google Scholar]
- Arpaia N, Green JA, Moltedo B, et al. A Distinct function of regulatory T cells in tissue protection. Cell. 2015 ; 162 : 1078–1089. [CrossRef] [PubMed] [Google Scholar]
- Gieseck RL, 3rd, Wilson MS, Wynn TA. Type 2 immunity in tissue repair and fibrosis. Nat Rev Immunol. 2018 ; 18 : 62–76. [CrossRef] [PubMed] [Google Scholar]
- de Kleer IM, Kool M, de Bruijn MJ, et al. Perinatal activation of the interleukin-33 pathway promotes type 2 immunity in the developing lung. Immunity. 2016 ; 45 : 1285–1298. [CrossRef] [PubMed] [Google Scholar]
- Saluzzo S, Gorki AD, Rana BMJ, et al. First-breath-induced type 2 pathways shape the lung immune environment. Cell rep. 2017 ; 18 : 1893–1905. [CrossRef] [PubMed] [Google Scholar]
- Ricardo-Gonzalez RR, Van Dyken SJ, Schneider C, et al. Tissue signals imprint ILC2 identity with anticipatory function. Nat Immunol. 2018 ; 19 : 1093–1099. [CrossRef] [PubMed] [Google Scholar]
- Cohen M, Giladi A, Gorki AD, et al. Lung single-cell signaling interaction map reveals basophil role in macrophage imprinting. Cell. 2018 ; 175 : 1031–44. e18. [CrossRef] [PubMed] [Google Scholar]
- Vasanthakumar A, Moro K, Xin A, et al. The transcriptional regulators IRF4, BATF and IL-33 orchestrate development and maintenance of adipose tissue-resident regulatory T cells. Nat Immunol. 2015 ; 16 : 276–285. [CrossRef] [PubMed] [Google Scholar]
- Bessa J, Meyer CA, de Vera Mudry MC, et al. Altered subcellular localization of IL-33 leads to non-resolving lethal inflammation. J Autoimmunity. 2014 ; 55 : 33–41. [CrossRef] [Google Scholar]
- Gautier V, Cayrol C, Farache D, et al. Extracellular IL-33 cytokine, but not endogenous nuclear IL-33, regulates protein expression in endothelial cells. Sci Rep. 2016 ; 6 : 34255. [CrossRef] [PubMed] [Google Scholar]
- Cohen ES, Scott IC, Majithiya JB, et al. Oxidation of the alarmin IL-33 regulates ST2-dependent inflammation. Nat comm. 2015 ; 6 : 8327. [CrossRef] [Google Scholar]
- Moffatt MF, Gut IG, Demenais F, et al. A large-scale, consortium-based genomewide association study of asthma. N Engl J Med. 2010 ; 363 : 1211–1221. [Google Scholar]
- Cayrol C, Girard JP. IL-33: an alarmin cytokine with crucial roles in innate immunity, inflammation and allergy. Cur Opin Immunol. 2014 ; 31 : 31–37. [CrossRef] [Google Scholar]
- Smith D, Helgason H, Sulem P, et al. A rare IL33 loss-of-function mutation reduces blood eosinophil counts and protects from asthma. PLoS Genet. 2017 ; 13 : e1006659. [PubMed] [Google Scholar]
- Kim YH, Yang TY, Park CS, et al. Anti-IL-33 antibody has a therapeutic effect in a murine model of allergic rhinitis. Allergy. 2012 ; 67 : 183–190. [CrossRef] [PubMed] [Google Scholar]
Liste des tableaux
Exemples de l’implication de l’axe IL-33/ST2 dans les maladies [12]. IL-33 : interleukine-33 ; ILC2 : cellules lymphoïdes innées de type 2 ; ST2 : suppression of tumorigenicity 2 ; sST2 : ST2 soluble ; NK : natural killer ; LT CD8+ : lymphocyte T CD8+ ; Th1 : lymphocyte T helper 1 ; DC : cellules dendritiques ; Th2 : lymphocyte T helper 2, Il1rl1 : Interleukin 1 receptor-like 1 ; IL-13 : interleukine 13 ; BPCO : bronchopneumopathie chronique obstructive ; DSS : dextran sodium sulfate ; TNBS : trinitrobenzene sulfonic acid ; Treg : lymphocytes T régulateurs ; RGCH : réaction du greffon contre l’hôte.
Anticorps monoclonaux thérapeutiques dans les maladies allergiques. Moduler les acteurs majeurs des réponses de type 2 représente une stratégie prometteuse pour traiter les maladies allergiques. Des thérapies à base d’anticorps visant à bloquer les IgE ou les cytokines de type 2 ont déjà été utilisées durant cette dernière décennie pour lutter contre l’asthme sévère et la dermatite atopique. Plusieurs grandes sociétés pharmaceutiques développent maintenant des anticorps monoclonaux ciblant directement la voie IL-33/ST2 dans l’espoir d’améliorer les traitements existants et d’étendre le spectre des maladies concernées. Les premiers résultats avec ces anticorps anti-IL-33/ST2, actuellement en essai clinique de phase I ou II, sont très encourageants. FDA : food and drug administration ; AMM : autorisation de mise sur le marché.
Liste des figures
|
Figure 1. Voies de signalisation de l’IL-33. Le récepteur ST2L (suppressor of tumorigenicity 2) comprend une partie extracellulaire composée de trois domaines Ig-like, un domaine transmembranaire et un domaine cytoplasmique TIR (Toll-likeinterleukin 1 receptor). La liaison de l’IL-33 à son récepteur ST2L entraîne un changement conformationnel qui permet la formation d’un complexe trimérique de haute affinité avec le corécepteur IL1RAcP (IL-1 receptor accessory protein), également corécepteur de l’IL-1 (interleukine 1) [4]. Comme pour l’IL-1, le rapprochement des deux domaines TIR entraîne le recrutement de la protéine adaptatrice MyD88 (myeloid differentiation primary response protein 88) qui initie la cascade de signalisation impliquant les molécules adaptatrices IRAK1 (IL-1b receptor-associated kinase 1), IRAK4 (IL-1b receptor-associated kinase 4), et TRAF6 (TNF receptor associated factor 6) [2]. Les voies IKK (IκB kinase)/NF-κB (nuclear factor-kappa B) et MAPK (mitogen-activated protein kinase) (p38, JNK [c-Jun N-terminal kinase] ou ERK [extracellular signal-regulated kinase]) sont alors activées, permettant notamment la translocation des facteurs de transcription AP-1 (activator protein 1) et NF-κB et la transcription de gènes cibles. |
| Dans le texte | |
|
Figure 2. Rôles de l’IL- 33 dans l’initiation des réponses immunes de type 2, au niveau des poumons. Suite à divers stimulus qui endommagent la barrière épithéliale du poumon (allergène, lésion, parasite, stress, etc.), l’IL-33 est libérée des cellules épithéliales. Très rapidement, elle active et induit la prolifération des cellules résidentes ILC2 (type 2 innate lymphoid cells), mastocytes et basophiles. Ces cellules sécrètent des cytokines Th2 (IL-4, IL-5, IL-13, IL-9) et divers médiateurs inflammatoires qui permettent la mise en place de la réponse innée de type 2. L’IL-13 favorise la migration des cellules dendritiques (DC) dans les noeuds lymphatiques qui vont permettre la différenciation des lymphocytes T naïfs (T Naïfs) en lymphocyte T helper 2 (Th2). L’IL-13 participe au recrutement, à l’activation et à la polarisation des macrophages alvéolaires M2. L’IL-5 est une cytokine clé pour le recrutement, l’activation et la survie des éosinophiles. L’ensemble de ces signaux conduit à l’inflammation des voies respiratoires, avec l’hyperplasie des cellules caliciformes, la production de mucus (qui permet l’expulsion des vers), l’hyperréactivité pulmonaire, l’éosinophilie et la production d’IgE (immunoglobuline E). Certains des signaux émis (amphiréguline ou AREG) permettent de réparer les lésions et participent au remodelage de l’épithélium. B : lymphocyte B ; Treg : lymphocyte T régulateur ; IL : interleukine. |
| Dans le texte | |
|
Figure 3. Les multiples rôles de l’IL-33 dans le système immunitaire. L’IL-33 est libérée des cellules productrices, majoritairement des cellules épithéliales et endothéliales, dans le milieu extracellulaire par divers stimuli (parasite, virus, allergène, lésion, etc.). Elle active et induit la prolifération ou permet la polarisation de nombreuses cellules cibles qui expriment le récepteur ST2L dans un contexte donné. L’ensemble des médiateurs sécrétés (cytokines, chimiokines ou autre) et les différentes interactions cellulaires qui sont alors établies vont permettre d’orienter la réponse immune. Ainsi, selon le tissu et l’environnement, l’IL-33 peut jouer des rôles très différents, et être impliquée dans des réponses pro- ou anti-inflammatoires, dans l’immunité antiparasitaire, antivirale, dans les processus de réparation tissulaire, etc. Des études récentes ont montré que l’IL-33 participait également à la genèse et/ou au maintien de l’homéostasie de différents tissus [38-40]. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.