")
")
| Issue |
Med Sci (Paris)
Volume 28, Number 2, Février 2012
|
|
|---|---|---|
| Page(s) | 185 - 192 | |
| Section | M/S Revues | |
| DOI | https://doi.org/10.1051/medsci/2012282017 | |
| Published online | 27 février 2012 | |
Le dimorphisme sexuel au XXIe siècle
Sexual dimorphism in the XXIst century
1
INRA, UMR1198, biologie du développement et reproduction, 78352 Jouy-en-Josas, France
2
École nationale vétérinaire d’Alfort, unité de biologie du développement et reproduction, F-94704 Maisons Alfort, France
3
université de Versailles Saint-Quentin-en-Yvelines, domaine de Vilvert, bâtiment 230 F-78352 Jouy-en-Josas, France
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
**
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
***
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Résumé
En ce début du XXIe siècle, une redéfinition du dimorphisme sexuel s’impose. Elle se doit d’incorporer conjointement non seulement les hormones sexuelles et le formatage socioculturel spécifique du genre, mais aussi l’importance des gènes localisés sur les chromosomes sexuels. Ces différentes composantes ont des effets indépendants et parallèles et qui interagissent dès la conception et tout au long de la vie. Des mécanismes épigénétiques assurent la mise en place de marques spécifiques du sexe qui modulent l’expression des gènes sans changer leur séquence. Ces marques représentent une sorte de mémoire pour se « souvenir » de son sexe, mais aussi pour « archiver » les impacts de l’environnement, selon l’expérience. Dans tous les tissus, ces marques et d’autres à venir, façonnées en fonction du sexe et du genre au gré de l’environnement, établissent des réseaux de gènes différents chez le mâle et la femelle, tant au niveau basal que pour les réponses immédiates et futures.
Abstract
A new definition of sexual dimorphism is required. The divergent biology of the sexes is still largely ignored, overshadowed by sociocultural considerations and confined to its hormonal organizational and activational effects, while the genes unequally expressed by the sex chromosomes play an important role much earlier, after conception, to set the stage and throughout life. These different components have independent and parallel effects that can interact in a synergistic or antagonistic manner on differentiation and response processes to trigger or erase sex-specific differences. The epigenetic marks and machinery represent the perfect tools to keep the memory of which sex is ours from the very beginning of life. Within the context of the developmental origin of adult health and diseases (DOHaD), owing to their flexibility to the environment, epigenetic marks also represent a support to archive the effects of environments during development, according to the sex of the parent, in a sex-specific mode. In all tissues, including gonads and brain, different trajectories of genes and pathways are used at the basal levels and to modulate/dictate responses according to sex and gender. It is urgent to emphasize the need to take into consideration this new knowledge and to apply less sex-biased approaches in research, medicine and society, to enhance women health and well-being. A critical review and realization of gender-specific social constraints, an indeniably but slowly on-going process, should allow us to “set free our sex biology” while detracting the delusion of hierarchy of the complex mechanisms involved.
© 2012 médecine/sciences – Inserm / SRMS
« If not for sex, much of what is flamboyant and beautiful in nature would not exist. Plants would not bloom. Birds would not sing. Deers would not sprout antlers. Hearts would not beat so fast. » (Dr Tatiana in [1]).
Les composantes du dimorphisme sexuel
Dès l’instant de notre conception, chacun d’entre nous est doté d’un sexe biologique [2]. Sur les 23 paires de chromosomes héritées de nos parents, c’est la paire de chromosomes sexuels qui fait toute la différence, XX pour une fille et XY pour un garçon. Il n’existe que de très rares cas (0,018 % des naissances) pour lesquels le sexe, à la naissance, peut être ambigu, que l’origine soit chromosomique, génique, développementale ou environnementale [3].
Bien que le sexe soit considéré comme déterminé essentiellement, et initialement, de manière biologique, notre genre (c’est-à-dire la perception sociale et les implications de notre sexe) est sans aucun doute au moins, si ce n’est plus, important pour nos vies. Les différences liées au sexe et celles liées au genre sont créées par l’intrication réciproque de multiples forces biologiques et environnementales [5]. Celles qui sont liées au sexe - qu’il s’agisse de l’âge d’apparition, de la sévérité ou de la prévalence de nombreuses maladies, du métabolisme, de la réponse aux médicaments ou aux régimes, de comportements, mais aussi de différences dans la programmation au cours du développement - sont largement reconnues. Une large part des études sur l’origine développementale de la santé et des maladies de l’adulte (DOHaD pour International society for developmental origins of health and disease1) fait état des modes de transmission dus au sexe et/ou des effets spécifiques du sexe, celui du parent et celui de la progéniture [4].
En dépit de ces différences, comme un éditorial de la revue Nature l’a récemment souligné2, la parité en recherche biomédicale n’est toujours pas atteinte. La majorité des études s’adressent à un seul sexe – généralement les mâles – aussi bien pour les études chez l’animal que pour les études cliniques chez l’homme. Si la médecine appliquée aux hommes est basée sur l’évidence scientifique, celle qui s’adresse aujourd’hui aux femmes l’est beaucoup moins. Au-delà des aspects éthiques interdisant le plus souvent les essais cliniques pendant la période de gestation/lactation (voir à ce sujet l’article de F. Nosten publié dans médecine/sciences [42]), et du fait de soins et de pratiques calqués sans discernement sur ceux qui sont appliqués aux hommes, l’effet, non intentionnel, est de négliger la santé des femmes [5].
La biologie du sexe, qui sous-tend ces différences, a longtemps été occultée par des considérations socioculturelles et confinée à l’étude du rôle des hormones alors que le sexe chromosomique joue aussi un rôle important et beaucoup plus précocement (Figure 1). Des différences sont en effet générées chez les invertébrés en l’absence d’hormones sexuelles : elles doivent donc trouver leur origine dans les chromosomes sexuels de ces espèces. En revanche chez les vertébrés, ces différences étaient communément attribuées aux seuls effets des hormones, mais des études sur des modèles de mammifères et d’oiseaux ont remis en cause cette explication [6]. Il est en effet bien connu que l’ablation hormonale ne parvient pas toujours à éliminer complètement ces différences, ni la supplémentation hormonale à les récréer [7]. Le rôle des chromosomes sexuels [8] doit désormais être pris en compte, au même titre que celui des hormones et des influences socioculturelles, pour rendre compte de ces différences (Figure 2).
 |
Figure 1. Les gènes des chromosomes sexuels ne sont pas tous équivalents et peuvent s’exprimer différemment selon le sexe de l’individu. Les cellules mâles possèdent un seul chromosome X d’origine maternelle et un chromosome Y d’origine paternelle. Les cellules femelles ont deux chromosomes X, mais un seul est actif : le chromosome X actif est soit d’origine maternelle (Xma, rectangle rouge), soit d’origine paternelle (Xpa, rectangle bleu). L’expression tissulaire est la résultante de l’expression de ces deux types de cellules. L’inactivation au hasard, entraînant une mosaïque, explique que certains traits, comme des traits cognitifs, montrent un certain degré de variabilité plus important chez les femelles que chez les mâles. La région pseudo-autosomique (PAR rectangle blanc) est commune à l’X et à l’Y et échappe à l’inactivation de l’X. Plusieurs classes de gènes peuvent s’exprimer de manière sexuellement dimorphique, selon leur position et leur statut sur les chromosomes X et Y : (1) les gènes spécifiques de l’Y sont exprimés uniquement chez le mâle (flèche bleue) ; (2) les gènes qui échappent à l’inactivation de l’X (flèches rouges) et ont un homologue différent sur le chromosome Y, fonctionnel ou non (flèche rose sur le Y), seront plus fortement exprimés chez la femelle ; (3) les gènes soumis à empreinte exprimés par le chromosome maternel/portés par le chromosome X (flèches blanches) et qui échappent à l’inactivation de l’X seront plus fortement exprimés chez le mâle que chez la femelle ; (4) les gènes soumis à empreinte exprimés par le chromosome paternel (échappant ou non à l’inactivation) sont exprimés uniquement chez la femelle (flèches orange et violette). À l’inverse, d’autres catégories de gènes peuvent s’exprimer de façon équivalente chez le mâle et chez la femelle. Ces catégories incluent les gènes soumis à l’inactivation de l’X, les gènes soumis à empreinte exprimés par le chromosome maternel/portés par le chromosome X et soumis à l’inactivation de l’X (flèches vertes) et les gènes de la région pseudo-autosomique (PAR) (flèches noires), qui est commune à l’X et à l’Y et échappent à l’inactivation de l’X, et les gènes liés à l’X qui s’expriment à partir de l’allèle maternel et échappent à l’inactivation de l’X (flèches jaunes). L’expression génique, à la fois dans les cellules mâles et les cellules femelles, peut être influencée, parfois de manière spécifique du sexe, par des facteurs externes incluant les influences sociales et le milieu hormonal [8]. |
 |
Figure 2. Facteurs génétiques et de proximité entraînant des différences phénotypiques liées au sexe. Le chromosome X comprend environ 1 500 et le chromosome Y 78 gènes. Certains de ces gènes s’expriment différemment selon qu’il s’agit d’un mâle (46 XY) ou d’une femelle (46 XX). Certains aspects différents du métabolisme et de la croissance peuvent être attribués à l’expression déséquilibrée des gènes liés à l’X entre les sexes, en particulier à certains stades du développement préimplantatoire, quand les 2 chromosomes X peuvent être actifs. Si l’on considère l’organisme entier, les chromosomes sexuels jouent un rôle crucial dans l’établissement du dimorphisme sexuel des fonctions cellulaires. Mais cette expression déséquilibrée n’est pas confinée au développement préimplantatoire, elle intervient aussi ultérieurement, puisque certains gènes portés par le chromosome X échappent à l’inactivation ou sont soumis à empreinte. L’initiation de la différenciation sexuelle dépend du gène SRY, codé par le chromosome Y. Les gènes de détermination du sexe situés sur les chromosomes sexuels peuvent influencer non seulement le développement et le maintien différencié d’organes sexuels secondaires non gonadiques, mais aussi ceux d’organes en dehors du système reproductif, comme le foie, le cerveau [40]. Les hormones ont deux types d’effets : permanents « organisationnels » sur les organes génitaux, le cerveau et peut-être d’autres tissus, et des effets aigus « d’activation », via les hormones ovariennes et testiculaires. L’environnement, sous toutes ses formes, intervient à tous les stades avant et pendant la gestation et après la naissance, via des effets biologiques liés au sexe ou des aspects à proprement parler socioculturels contribuant à sculpter le phénotype social basé sur le genre. Toutes ces composantes envoient des signaux à la machinerie et aux marques épigénétiques, permettant à l’épigénome de chaque cellule de refléter le sexe (le genre) de l’individu (flèches vertes). |
Très peu d’études encore se sont attachées à décrire les mécanismes intervenant aux niveaux transcriptionnel et épigénétique, et encore moins ceux qui résultent d’influences environnementales, nutritionnelles ou sociétales.
L’objectif de cet article est de décrire les connaissances récemment acquises et de montrer comment le génome des organes, au cours du développement puis à l’âge adulte, se façonne au gré de l’environnement et retient la mémoire des événements passés longtemps après l’impact de l’exposition - en fonction du sexe biologique, puis du genre.
Importance des niveaux du dimorphisme sexuel aux différents stades de la vie
Le dimorphisme sexuel a été étudié à divers stades : de la conception jusqu’aux stades précédant la différenciation gonadique, au cours du développement (et plus particulièrement dans l’embryon et le placenta) et enfin chez l’adulte, surtout dans le cerveau et le foie.
Stades précoces précédant la différenciation gonadique
Les signaux de proximité qui agissent directement sur les cellules pour provoquer la différenciation sexuelle ne se résument pas aux hormones gonadiques, mais incluent d’autres facteurs comme les produits des gènes portés par les chromosomes sexuels et ceux de gènes non gonadiques régulés en aval (pour revue [4]) (Figure 2).
Dans un blastocyste bovin, un dimorphisme sexuel influence l’expression d’un tiers des gènes qui s’expriment [9]. Dans le blastocyste de souris également, près de 600 gènes sont différentiellement exprimés entre le mâle et la femelle [10]. Ceci démontre qu’en l’absence d’influences hormonales à ce stade, les chromosomes sexuels peuvent imposer une régulation transcriptionnelle à des gènes autosomiques. L’analyse ontologique des données chez le bovin suggère un niveau transcriptionnel global plus important chez les femelles et un métabolisme protéique plus important chez les mâles. D’un point de vue phénotypique, le métabolisme global du glucose est deux fois plus important et l’activité de la voie des pentoses phosphates quatre fois plus importante dans les blastocystes bovins mâles que dans les blastocystes femelles. Des différences métaboliques similaires ont été observées chez les embryons humains au même stade (pour revue, voir [11]).
Bien avant la différenciation des gonades, un dimorphisme sexuel existe entre des cellules embryonnaires mâles et des cellules femelles vis-à-vis de leur sensibilité (plus importante chez les femelles) à une exposition à l’alcool, ce qui peut être attribué à l’action de gènes portés par les chromosomes sexuels [12].
On sait également que les différences de développement du cerveau et de comportement entre les mâles et les femelles, aux stades précédant la différenciation gonadique, résultent directement des profils d’expression sexuellement dimorphiques de gènes liés au développement neuronal. Au jour 10,5 du développement embryonnaire chez la souris, on peut détecter l’expression différentielle d’une cinquantaine de gènes, pour la plupart autosomiques, entre les cerveaux d’embryons mâles et femelles [13]. Dans leur très grande majorité, ces gènes interviennent dans trois catégories de fonctions biologiques : différenciation et prolifération cellulaires, régulation transcriptionnelle et signalisation. On trouve également des gènes codant pour des enzymes de la machinerie épigénétique. Citons Jarid1c et Jarid1d - déméthylases de la lysine 4 de l’histone H3 - portés respectivement par l’X et l’Y-, et Utx et son homologue Uty, correspondant à une déméthylase de la lysine 27 de l’histone H3.
Les mécanismes par lesquels ces gènes agissent plus tard sur le développement des neurones et le comportement, indépendamment ou en synergie avec les hormones gonadiques, restent à déterminer.
Ainsi, du fait de l’expression déséquilibrée des gènes liés à l’X entre les sexes à certains stades du développement préimplantatoire précoce, le sexe de l’embryon joue un rôle déterminant dans l’établissement de la taille de l’embryon, sa vitesse de croissance, le métabolisme, la réponse et la morbidité face à certains stimulus délétères (Figure 1).
Périodes fœtale et périnatale : l’exemple du dimorphisme sexuel dans le développement cérébral
Pendant longtemps les tissus extraembryonnaires, en particulier le placenta, ont été considérés comme des organes asexués, la plupart des études ne prenant pas en compte le sexe de l’embryon [14]. Des travaux récents - dont ceux de notre laboratoire - ont montré, par une approche d’analyse de gènes candidats ou transcriptomique, des profils d’expression sexuellement dimorphiques dans le placenta de bovins, de rongeurs et humains [14–17]. En réponse à différents stimulus nutritionnels, par exemple le régime alimentaire maternel, nous avons identifié une réponse différente des placentas mâles et femelles [18]. Dans le cas de l’asthme maternel, par exemple, les croissances fœtale et placentaire sont très différentes entre les garçons et les filles [14]. Les effets inflammatoires de la maladie maternelle affectent la croissance de la femelle ; à l’inverse, le fœtus mâle continue à croître normalement [8], et n’est affecté que si la maladie s’exacerbe de façon aiguë [14].
On a longtemps pensé que la différenciation sexuelle du cerveau - organe « bipotentiel » - et celle du comportement, étaient directement influencées par l’action des hormones gonadiques stéroïdes. Les changements des taux d’hormones gonadiques au cours de la vie et d’autres changements dynamiques conditionnent les différences liées au sexe et les accentuent ou les suppriment au cours du temps. Parmi les nombreuses différences liées au sexe, citons par exemple : le volume des noyaux de certaines régions cérébrales, le nombre des neurones, la densité et le nombre des synapses, la morphologie gliale [19]. Des régions distinctes de l’hypothalamus se réorganisent différemment sous l’influence de la même hormone stéroïde, et cette spécificité de la différenciation sexuelle cellulaire a été bien caractérisée dans trois noyaux hypothalamiques : le noyau arqué, la région préoptique et l’hypothalamus médian basal. Le volume du noyau sexuellement dimorphique de l’aire médiane préoptique (critique pour l’expression du comportement sexuel du mâle) est cinq fois plus grand chez les mâles. C’est l’inverse pour le noyau antéroventral périventriculaire. De subtiles différences liées au sexe ont été identifiées dans l’hippocampe ; elles seraient liées à des comportements différents dans les stratégies d’apprentissage spatial, la réponse au stress, et l’impact à long terme d’événements précoces néfastes (pour revue, voir [2]). Dans des régions spécifiques du cerveau, les hormones testiculaires influencent la mort et la prolifération cellulaires au cours du développement périnatal, dans un sens ou dans l’autre [19]. Par exemple, un traitement par l’œstradiol exogène promeut la prolifération cellulaire et la survie de l’hippocampe chez la femelle en période néonatale mais pas chez le mâle. Un antagoniste de l’œstradiol endogène entraîne une réduction de la prolifération cellulaire dans l’hippocampe du mâle mais pas dans celui de la femelle.
Des données récentes basées sur l’imagerie, notamment l’IRM (imagerie à résonance magnétique), ont effectivement montré que de nombreuses structures du cerveau se développent et/ou fonctionnent d’une manière sexuellement dimorphique [20]. C’est particulièrement évident au niveau du cortex : le volume des aires corticales frontale et médiane paralimbique est plus important chez les femmes, et celui du cortex fronto médian, de l’amygdale et de l’hypothalamus chez l’homme. Ces différences peuvent contribuer aux divergences entre mâles et femelles dans la réalisation de tâches ou même expliquer certains comportements stéréotypiques différents [5, 20]. Il a été suggéré que l’impact des aggressions précoces prédisposant à la schizophrénie pouvait avoir des conséquences différentes sur les régions du cerveau structurellement différentes chez l’homme ou la femme (pour revue, voir [20]).
Influences socioculturelles spécifiques du genre après la naissance
Enfin, dès la naissance, les aspects socioculturels prennent le pas et ont indéniablement un effet considérable : ils sculptent de façon très efficace le phénotype social basé sur le genre. On a longtemps pensé que ces effets étaient limités à l’espèce humaine.
Pourtant il existe une abondante littérature, surtout chez le rat, montrant que, dès la première semaine de vie, les mères se comportent différemment selon le sexe de leur progéniture. Ainsi, la mère consacre deux fois plus de temps aux soins de la région anogénitale des petits mâles par rapport aux petits de sexe féminin. Cette stimulation somatosensorielle est nécessaire au développement des nerfs contrôlant le pénis.
Le degré d’attention de la mère induit une différence de méthylation du promoteur du gène codant pour le récepteur des œstrogènes dans l’aire médiane préoptique ; celle-ci entraîne une différence dans l’expression de ce gène plus tard au cours du développement. Ainsi est induit chez la femelle un comportement comparable à celui de sa mère : une mère « plus attentionnée » donnera des filles qui deviendront à leur tour des mères « plus attentionnées » et réciproquement. Inversement une diminution des soins maternels augmente l’expression de ce récepteur dans d’autres régions, comme le noyau antéroventral périventriculaire de l’hypothalamus, plus développé chez la femelle, et le noyau ventromédian de l’hypothalamus, deux aires critiques pour le contrôle de la sécrétion de gonadotrophines, du cycle œstral, et du comportement sexuel de la femelle. Il existe aussi une relation entre ce comportement maternel et un important dimorphisme sexuel dans différents comportements de la progéniture, par exemple dans la durée du jeu chez les mâles [2, 21].
Mais peut-on pour autant qualifier cette pratique biologique, basée sur la reconnaissance de signaux urinaires, de socioculturelle ? Il y a toujours eu une certaine réticence à admettre chez l’homme l’existence de mécanismes analogues basés sur des phéromones. Et pourtant, un père, une mère ne traitent-ils pas différemment un garçon et une fille, et ce dès leur naissance ? Pourquoi n’existerait-il pas, chez l’homme aussi, des bases biologiques analogues ? Il sera toutefois difficile de faire la différence entre les pratiques socioculturelles et des mécanismes biologiques (Figure 2).
Dimorphisme sexuel chez l’adulte
Les mécanismes qui régulent les différences entre les sexes ont été très étudiés dans le foie de rongeurs, mais tendent à l’être dans d’autres organes murins, comme le rein, les glandes lacrymales, le cerveau ou le cœur [4, 24]. Un dimorphisme sexuel existe probablement pour tous les tissus, mais s’exprime dans des proportions qui varient selon l’âge et les conditions physiopathologiques et environnementales.
Une analyse transcriptomique faite sur du tissu musculaire, hépatique et cérébral a révélé que l’importance du dimorphisme sexuel dans l’expression des gènes est beaucoup plus grande qu’on ne l’imaginait jusqu’alors [24]. D’après ce travail, le dimorphisme sexuel concerne 14 % des gènes dans le cerveau total et 70 % dans le foie, avec une forte spécificité des profils d’expression selon le tissu considéré, mais la proportion de gènes sexuellement dimorphiques communs aux différents tissus est relativement faible (8 %). Ces gènes sont regroupés non seulement sur les chromosomes sexuels mais aussi sur les autosomes, suggérant des mécanismes de corégulation [24].
Dans le tissu adipeux, le cerveau, le foie et le muscle, ce dimorphisme sexuel est détecté à l’échelle de modules de réseaux régulés par ces loci chromosomiques et par les hormones gonadiques. Ces réseaux spécifiques du sexe sont fortement corrélés à des traits génétiques et métaboliques [25]. C’est le cas pour les gènes spécifiquement mâles comme Cyp2a2, ou femelles comme Cyp2c12, codant pour des cytochromes P450 (voir ci-dessous) [25]. Dans une autre étude portant sur le foie adulte, on a pu estimer que la part du rôle des gènes localisés sur les chromosomes sexuels dans la modulation de l’expression spécifique du sexe semble modeste (10 %), par rapport au rôle dominant des hormones sexuelles [26]. Il reste à déterminer les parts respectives de ces composantes dans le déterminisme, le maintien et l’évolution du dimorphisme sexuel en fonction du sexe, du stade de développement, du tissu, de l’âge, de l’état physiopathologique de la réponse à l’environnement et de l’expérience.
On rencontre aussi de telles différences liées au sexe dans les domaines de la pharmacocinétique et de la pharmacodynamie, et elles contribuent aux différences individuelles dans la réponse, l’efficacité et la toxicité des médicaments. Ces différences s’expliquent par la variation d’expression des enzymes hépatiques telles que les cytochromes P450 impliqués dans le métabolisme des médicaments, des stéroïdes, des acides gras et des composés chimiques environnementaux. Chez le rat, l’expression sexuellement dimorphique des P450 et des autres gènes spécifiquement exprimés dans le foie est régulée par les différences de profils temporels de l’hormone de croissance plasmatique (GH, growth hormone), dont la sécrétion est pulsatile chez le mâle et continue chez la femelle [27]. Des différences analogues existent également chez l’homme [27].
Les mécanismes et les marques épigénétiques : l’interface entre gènes et environnement
Comment le génome des organes en cours de développement et chez l’adulte peut-il se « souvenir » de son sexe biologique qui s’est imprimé progressivement depuis la conception ? Les marques épigénétiques sont apposées sur la séquence de l’ADN (méthylation de l’ADN) ou portées par les protéines (histones) auxquelles il s’associe. Ces processus, qui interviennent dans tous les mécanismes vitaux (transcription, réparation, réplication, condensation, inactivation d’un chromosome X, empreinte parentale, vieillissement), sont essentiels pour le développement, la prolifération et la différenciation cellulaires chez tous les êtres vivants [28]. Un remodelage de la chromatine et des marques laissées par la machinerie épigénétique au cœur de complexes protéiques se produit également tout au long de la journée et de la vie sous l’influence des horloges biologiques (circadiennes, saisonnières), des environnements complexes (nutritionnels, sociaux) auxquels nous sommes soumis, de nos expériences et de notre vécu. Grâce à leur flexibilité, ces mécanismes et les marques épigénétiques constituent l’interface entre les gènes - immuables dans leur séquence - et l’environnement, qui fluctue en permanence (revue dans [29]). Ces marques constituent un mode d’archivage privilégié pour stocker, selon un mode qui est spécifique au sexe, la mémoire des événements, des adaptations, des apprentissages, des expériences passées qu’ils soient de nature chimique (métabolique, nutriments, toxiques) [30] ou autre (social, affectif, exercice, psychobiologique) [23]. Elles pourront ainsi contrôler l’expression de jeux de gènes clés pour moduler les réponses immédiates et futures de l’individu [31].
Ainsi l’établissement, dès les stades les plus précoces du développement, et la conservation de différences liées au sexe pourraient être contrôlés par des marques épigénétiques spécifiques du sexe. Dans les blastocystes bovins, l’expression d’enzymes clés de la machinerie épigénétique, comme les ADN méthyltransférases Dnmt1 (augmentée chez les femelles), Dnmt3a et Dnmt3b ou l’histone méthyltransférase Hmt1 (toutes diminuées chez les femelles), est dépendante du sexe [32]. Ces différences épigénétiques suggèrent ainsi que, bien avant la différenciation des gonades, des mécanismes épigénétiques peuvent être impliqués dans les différences dans la vitesse de développement, le métabolisme et la transcription décrites précédemment (Figure 2).
Des altérations dans la programmation épigénétique des différences liées au sexe dans le cerveau pourraient rendre compte du dimorphisme sexuel des affections neurodéveloppementales [33]. Un dimorphisme sexuel a été mis en évidence pour l’expression de Dnmt3a, du gène codant la protéine de liaison à l’ADN méthylé MeCP2, et du nuclear corepressor Ncor, et pour les profils de méthylation de l’ADN dans le cerveau de rongeur en développement. Ces différences ont un effet à court et long terme sur le comportement social des rats en matières de jeu et d’anxiété [34]. Les perturbations exogènes ou endogènes au cours du développement peuvent avoir un impact sur ces processus épigénétiques et donc des conséquences profondes sur l’expression génique. Altérant ainsi les processus de développement du cerveau, elles peuvent conférer un risque ou une résilience à un défaut neurologique ou une affection mentale, de façon spécifique du sexe [2, 23].
Des différences de structure de la chromatine et des marques épigénétiques entre mâles et femelles ont été rapportées pour différents tissus [17, 24, 33]. Des modifications post-traductionnelles de l’histone H3, acétylation et méthylation, sont sexuellement dimorphiques dans le cerveau de souris en développement. Alors que le traitement par la testostérone in utero modifie les profils d’acétylation des embryons femelles, les rendant plus proches de ceux des mâles, il n’a pas d’effet sur la méthylation de l’histone H3, pourtant sexuellement dimorphique. Le contrôle de cette marque est donc indépendant des hormones [35].
L’épigénome du cerveau, comme de tous les tissus, en permanente interaction avec son environnement, reste malléable tout au long de la vie, prêt à enregistrer, à recomposer ou à effacer ! Chez l’homme, on commence aussi à déchiffrer les bases biologiques épigénétiques des effets socioculturels précoces [22]. Mais ce sont surtout les travaux effectués chez le rongeur (effets épigénétiques du stress précoce, de différences de comportements maternels) qui montrent comment des mécanismes épigénétiques sont capables de garder la mémoire des effets environnementaux dans différentes régions du cerveau, tout au long de la vie [23].
Dans le foie adulte, il existe maintenant de solides arguments pour impliquer les caractéristiques conformationnelles de la chromatine dans la régulation des gènes montrant un dimorphisme sexuel. Des différences dans l’hypersensibilité à la DNaseI, témoin de l’accessibilité à la chromatine, indiquent une organisation de la chromatine spécifique du sexe dans le foie. Celle-ci est vraisemblablement établie et/ou maintenue par les profils sexuellement dimorphiques de la sécrétion de l’hormone de croissance (GH). La GH est sécrétée de manière pulsatile chez le mâle mais continue chez la femelle, ce qui influence l’activité du facteur de transcription nucléaire STAT5b qui module uniquement l’expression de gènes spécifiquement mâles [27, 36].
Les mécanismes épigénétiques représentent aussi un moyen pour l’individu de transmettre certains caractères acquis à sa descendance. En effet, il a été montré que la programmation in utero (blastocyste, fœtus, placenta, nouveau-né) est sexuellement dimorphique si l’on considère non seulement le sexe du conceptus, mais aussi le sexe du parent soumis à une exposition environnementale [4, 37] (Figure 3).
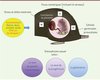 |
Figure 3. Dimorphisme sexuel des effets intergénérationnels et transgénérationnels. Le premier mode de transmission, de loin le mieux connu et étudié, est somatique. Il résulte de l’influence, sur les cellules somatiques du fœtus et du nouveau-né au cours du développement, de perturbations métaboliques ou nutritionnelles ou d’un stress de la mère pendant la grossesse et l’allaitement. Il peut se reproduire à chaque grossesse et engendrer des effets intergénérationnels. Le second mode de transmission passe par la lignée germinale. Alors que les marques épigénétiques des gamètes s’effacent, au cours de deux phases, la différenciation sexuelle et immédiatement après la fécondation, ces remises à zéro peuvent n’être pas complètes et épargner quelques marques ayant mémorisé le vécu d’un ancêtre. L’altération de certaines marques, non effacées, sur certains gènes dans les cellules germinales, pourrait ainsi être transmise à plusieurs générations successives (F1, F2 via les cellules germinales), sans toutefois que l’on en connaisse les mécanismes. Un dimorphisme sexuel peut dépendre du sexe du parent, de la progéniture et être différent aussi en fonction de la génération (revue dans [4, 41]). |
Conclusions et perspectives
Les organismes mâles et femelles ont donc des susceptibilités significativement différentes, mais les effets spécifiques du sexe sont encore trop souvent considérés comme facteur confondant plutôt que d’importance primordiale. Ces différences, très précoces, sont encore mal explorées mais devraient faire l’objet d’une attention plus soutenue, tant les stratégies thérapeutiques ou préventives qui sont efficaces pour les individus d’un sexe ne sont pas nécessairement adaptées à l’autre sexe [4, 29, 38, 39].
Les études qui, lors de leur conception ou de l’interprétation des résultats, ignorent ces données risquent de ne pas identifier une proportion importante de gènes qui contribuent au développement de maladies ou de comportements complexes [5]. Et même si les réponses semblent équivalentes, les trajectoires empruntées peuvent être différentes, et leur connaissance pourrait offrir de nouvelles cibles.
Des analyses bio-informatiques fondées sur la biologie des systèmes devraient permettre de déterminer s’il existe ou non des assortiments de gènes et des voies épigénétiques « universels » impliqués dans le dimorphisme sexuel et/ou si les bases du dimorphisme ont des spécificités spatiotemporelles communes aux différentes espèces. La connaissance des mécanismes de base du dimorphisme sexuel et de leurs effets très précoces, dès le stade blastocyste, voire dès le stade de démarrage de la transcription embryonnaire, permettra d’appréhender toute la diversité de nos capacités de réaction et d’adaptation à notre environnement nutritionnel, sociétal ou au stress et, peut-être, de « s’adapter » selon son sexe, voire son genre.
Nous nous devons d’inventorier et de déchiffrer les mécanismes par lesquels les gènes des chromosomes sexuels et les hormones sexuelles régulent les gènes de la machinerie épigénétique et contribuent aux différences liées au sexe via l’établissement d’épigénomes spécifiques d’un tissu, d’un stade, d’un état physiopathologique ou de l’âge. Il faudra décrypter la fonction biologique, et déterminer l’avantage ou le risque éventuel associé aux spécificités mâles et femelles.
La remise à plat du carcan des contraintes socioculturelles diverses s’impose si nous voulons accéder à nos aspirations légitimes, scientifiques médicales ou sociétales, ou, comme l’a écrit, à son époque, François Poullain de la Barre (1671) : « De l’égalité des deux sexes, discours physique et moral où l’on voit l’importance de se débarrasser des préjugés. » Depuis une trentaine d’années en effet, au nom de la parité entre les genres, on tendait à ignorer les différences entre les sexes. Les évidences scientifiques apportées aujourd’hui, en particulier au niveau moléculaire, attestent indéniablement de la réalité biologique du dimorphisme sexuel mais aussi de sa complexité. Il ne faut pas pour autant vouloir à tout prix attribuer à ces différences entre les sexes une justification évolutionniste, voire sociétale ou philosophique, ni surtout céder à la tentation - simpliste, au vu de leur complexité - de les hiérarchiser. Un gène, une cellule, un tissu, et, a fortiori un sexe, ne pense pas, n’a pas de dessein intelligent, et encore moins de finalité !
Conflits d’intérêts
Les auteurs déclarent n’avoir aucun conflit d’intérêts concernant les données publiées dans cet article
Remerciements
Claudine Junien remercie Jean-Claude Kaplan pour ses précieux conseils, ses encouragements, et son indéfectible soutien à la cause du sexe biologique.
Editorial : Putting gender on the agenda. Nature 2010 ; 465 : 665.
Références
- Williams TM, Carroll SB. Genetic and molecular insights into the development and evolution of sexual dimorphism. Nat Rev Genet 2009 ; 10 : 797–804. [CrossRef] [PubMed] [Google Scholar]
- McCarthy MM, Arnold AP. Reframing sexual differentiation of the brain. Nat Neurosci 2011 ; 14 : 677–683. [CrossRef] [PubMed] [Google Scholar]
- Sax L. How common is intersex? a response to Anne Fausto-Sterling. J Sex Res 2002 ; 39 : 174–178. [CrossRef] [PubMed] [Google Scholar]
- Gabory A, Attig L, Junien C. Sexual dimorphism in environmental epigenetic programming. Mol Cell Endocrinol 2009 ; 25 : 8–18. [CrossRef] [Google Scholar]
- Arnold AP. Promoting the understanding of sex differences to enhance equity and aexcellence in biomedical science. Biol Sex Differ 2011 ; 1 : 1. [CrossRef] [Google Scholar]
- Burgoyne PS, Thornhill AR, Boudrean SK, et al. The genetic basis of XX-XY differences present before gonadal sex differentiation in the mouse. Philos Trans R Soc Lond B Biol Sci 1995 ; 350 : 253–261. [CrossRef] [PubMed] [Google Scholar]
- Ceribelli A, Pino MS, Cecere FL. Gender differences : implications for clinical trials and practice. J Thorac Oncol 2007 ; 2 : S15–S18. [CrossRef] [PubMed] [Google Scholar]
- Davies W, Wilkinson LS. It is not all hormones : alternative explanations for sexual differentiation of the brain. Brain Res 2006 ; 1126 : 36–45. [CrossRef] [PubMed] [Google Scholar]
- Bermejo-Alvarez P, Rizos D, Rath D, et al. Sex determines the expression level of one third of the actively expressed genes in bovine blastocysts. Proc Natl Acad Sci USA 2010 ; 107 : 3394–3399. [CrossRef] [Google Scholar]
- Kobayashi S, Isotani A, Mise N, et al. Comparison of gene expression in male and female mouse blastocysts revealed imprinting of the X-linked gene, Rhox5/Pem, at preimplantation stages. Curr Biol 2006 ; 16 : 166–172. [CrossRef] [PubMed] [Google Scholar]
- Bermejo-Alvarez P, Rizos D, Lonergan P, Gutierrez-Adan A. Transcriptional sexual dimorphism during preimplantation embryo development and its consequences for developmental competence and adult health and disease. Reproduction 2011 ; 141 : 563–570. [CrossRef] [PubMed] [Google Scholar]
- Penaloza C, Estevez B, Orlanski S, et al. Sex of the cell dictates its response : differential gene expression and sensitivity to cell death inducing stress in male and female cells. FASEB J 2009 ; 23 : 1869–1879. [CrossRef] [PubMed] [Google Scholar]
- Dewing P, Shi T, Horvath S, Vilain E. Sexually dimorphic gene expression in mouse brain precedes gonadal differentiation. Brain Res Mol Brain Res 2003 ; 118 : 82–90. [CrossRef] [PubMed] [Google Scholar]
- Clifton VL. Sex the human placenta : mediating differential strategies of fetal growth and survival. Placenta 2010 ; 31 (suppl) : S33–S39. [CrossRef] [PubMed] [Google Scholar]
- Sood R, Zehnder JL, Druzin ML, Brown PO. Gene expression patterns in human placenta. Proc Natl Acad Sci USA 2006 ; 103 : 5478–5483. [CrossRef] [Google Scholar]
- Mao J, Zhang X, Sieli PT, et al. Contrasting effects of different maternal diets on sexually dimorphic gene expression in the murine placenta. Proc Natl Acad Sci USA 2010 ; 107 : 557–562. [Google Scholar]
- Gallou-Kabani C, Gabory A, Tost J, et al. Sex-and diet-specific changes of imprinted ggene expression and DNA methylation in mouse placenta under a high-fat diet. PLoS One 2010 ; 5 : e14398. [CrossRef] [PubMed] [Google Scholar]
- Gabory A, Ferry L, Fajardy I, et al. Female- and male-specific transcriptomic and epigenetic placental signatures in response to a maternal high-fat diet in mice. 2012 (soumis pour publication). [Google Scholar]
- Forger NG. Control of cell number in the sexually dimorphic brain and spinal cord. J Neuroendocrinol 2009 ; 21 : 393–399. [CrossRef] [PubMed] [Google Scholar]
- Cahill L. Why sex matters for neuroscience. Nat Rev Neurosci 2006 ; 7 : 477–484. [CrossRef] [PubMed] [Google Scholar]
- Moore CL, Power KL. Variation in maternal care and individual differences in play, exploration, and grooming of juvenile Norway rat offspring. Dev Psychobiol 1992 ; 25 : 165–182. [CrossRef] [PubMed] [Google Scholar]
- McGowan PO, Sasaki A, D’Alessio AC, et al. Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nat Neurosci 2009 ; 12 : 342–348. [CrossRef] [PubMed] [Google Scholar]
- Curley JP, Jensen CL, Mashoodh R, Champagne FA. Social influences on neurobiology and behavior : Epigenetic effects during development. Psychoneuroendocrinology 2010 ; 36 : 352–357. [CrossRef] [PubMed] [Google Scholar]
- Yang X, Schadt EE, Wang S, et al. Tissue-specific expression and regulation of sexually dimorphic genes in mice. Genome Res 2006 ; 16 : 995–1004. [CrossRef] [PubMed] [Google Scholar]
- Van Nas A, Guhathakurta D, Wang SS, et al. Elucidating the role of gonadal hormones in sexually dimorphic gene coexpression networks. Endocrinology 2009 ; 150 : 1235–1249. [CrossRef] [PubMed] [Google Scholar]
- Wauthier V, Sugathan A, Meyer RD, et al. Intrinsic sex differences in the early growth hormone responsiveness of sex-specific genes in mouse liver. Mol Endocrinol 2010 ; 24 : 667–678. [CrossRef] [PubMed] [Google Scholar]
- Waxman DJ, Holloway MG. Centennial perspective : sex differences in the expression of hepatic drug metabolizing enzymes. Mol Pharmacol 2009 ; 76 : 215–228. [CrossRef] [PubMed] [Google Scholar]
- Bourc’his D, Proudhon C. Sexual dimorphism in parental imprint ontogeny and contribution to embryonic development. Mol Cell Endocrinol 2008 ; 282 : 87–94. [CrossRef] [PubMed] [Google Scholar]
- Kaminsky Z, Wang SC, Petronis A. Complex disease, gender and epigenetics. Ann Med 2006 ; 38 : 530–544. [CrossRef] [PubMed] [Google Scholar]
- Csoka AB, Szyf M. Epigenetic side-effects of common pharmaceuticals : a potential new field in medicine and pharmacology. Med Hypotheses 2009 ; 73 : 770–780. [CrossRef] [PubMed] [Google Scholar]
- Gabory A, Attig L, Junien C. Developmental programming and epigenetics. Am J Clin Nutr 2011 ; 94 (suppl 6) : S1943–S1952. [Google Scholar]
- Bermejo-Alvarez P, Rizos D, Rath D, et al. Epigenetic differences between male and female bovine blastocysts produced in vitro. Physiol Genomics 2008 ; 32 : 264–272. [PubMed] [Google Scholar]
- McCarthy MM, Auger AP, Bale TL, et al. The epigenetics of sex differences in the brain. J Neurosci 2009 ; 29 : 12815–12823. [CrossRef] [PubMed] [Google Scholar]
- Jessen HM, Auger AP. Sex differences in epigenetic mechanisms may underlie risk and resilience for mental health disorders. Epigenetics 2011 ; 6 : 857–861. [CrossRef] [PubMed] [Google Scholar]
- Tsai HW, Grant PA, Rissman EF. Sex differences in histone modifications in the neonatal mouse brain. Epigenetics 2009 ; 4 : 47–53. [CrossRef] [PubMed] [Google Scholar]
- Ling G, Sugathan A, Mazor T, et al. Unbiased, genome-wide in vivo mapping of transcriptional regulatory elements reveals sex differences in chromatin structure associated with sex-specific liver gene expression. Mol Cell Biol 2010 ; 30 : 5531–5544. [CrossRef] [PubMed] [Google Scholar]
- Fauquier L. Quand l’environnement du père influence l’expression génique chez l’enfant. Med Sci (Paris) 2011 ; 27 : 453–455. [CrossRef] [EDP Sciences] [PubMed] [Google Scholar]
- Pennisi E. Sex and Social Structure. Science 2009 ; 326 : 518–519. [CrossRef] [PubMed] [Google Scholar]
- Shepard KN, Michopoulos V, Toufexis DJ, Wilson ME. Genetic, epigenetic and environmental impact on sex differences in social behavior. Physiol Behav 2009 ; 97 : 157–170. [Google Scholar]
- Schlessinger D, Garcia-Ortiz JE, Forabosco A, et al. Determination and stability of gonadal sex. J Androl 2011 ; 31 : 16–25. [CrossRef] [Google Scholar]
- Dunn GA, Morgan CP, Bale TL. Sex-specificity in transgenerational epigenetic programming. Horm Behav 2010 ; 59 : 290–295. [CrossRef] [PubMed] [Google Scholar]
- Nosten F. Paludisme et grossesse : un dilemme thérapeutique. Med Sci (Paris) 2009 ; 25 : 867–869. [CrossRef] [EDP Sciences] [PubMed] [Google Scholar]
Liste des figures
|
Figure 1. Les gènes des chromosomes sexuels ne sont pas tous équivalents et peuvent s’exprimer différemment selon le sexe de l’individu. Les cellules mâles possèdent un seul chromosome X d’origine maternelle et un chromosome Y d’origine paternelle. Les cellules femelles ont deux chromosomes X, mais un seul est actif : le chromosome X actif est soit d’origine maternelle (Xma, rectangle rouge), soit d’origine paternelle (Xpa, rectangle bleu). L’expression tissulaire est la résultante de l’expression de ces deux types de cellules. L’inactivation au hasard, entraînant une mosaïque, explique que certains traits, comme des traits cognitifs, montrent un certain degré de variabilité plus important chez les femelles que chez les mâles. La région pseudo-autosomique (PAR rectangle blanc) est commune à l’X et à l’Y et échappe à l’inactivation de l’X. Plusieurs classes de gènes peuvent s’exprimer de manière sexuellement dimorphique, selon leur position et leur statut sur les chromosomes X et Y : (1) les gènes spécifiques de l’Y sont exprimés uniquement chez le mâle (flèche bleue) ; (2) les gènes qui échappent à l’inactivation de l’X (flèches rouges) et ont un homologue différent sur le chromosome Y, fonctionnel ou non (flèche rose sur le Y), seront plus fortement exprimés chez la femelle ; (3) les gènes soumis à empreinte exprimés par le chromosome maternel/portés par le chromosome X (flèches blanches) et qui échappent à l’inactivation de l’X seront plus fortement exprimés chez le mâle que chez la femelle ; (4) les gènes soumis à empreinte exprimés par le chromosome paternel (échappant ou non à l’inactivation) sont exprimés uniquement chez la femelle (flèches orange et violette). À l’inverse, d’autres catégories de gènes peuvent s’exprimer de façon équivalente chez le mâle et chez la femelle. Ces catégories incluent les gènes soumis à l’inactivation de l’X, les gènes soumis à empreinte exprimés par le chromosome maternel/portés par le chromosome X et soumis à l’inactivation de l’X (flèches vertes) et les gènes de la région pseudo-autosomique (PAR) (flèches noires), qui est commune à l’X et à l’Y et échappent à l’inactivation de l’X, et les gènes liés à l’X qui s’expriment à partir de l’allèle maternel et échappent à l’inactivation de l’X (flèches jaunes). L’expression génique, à la fois dans les cellules mâles et les cellules femelles, peut être influencée, parfois de manière spécifique du sexe, par des facteurs externes incluant les influences sociales et le milieu hormonal [8]. |
| Dans le texte | |
|
Figure 2. Facteurs génétiques et de proximité entraînant des différences phénotypiques liées au sexe. Le chromosome X comprend environ 1 500 et le chromosome Y 78 gènes. Certains de ces gènes s’expriment différemment selon qu’il s’agit d’un mâle (46 XY) ou d’une femelle (46 XX). Certains aspects différents du métabolisme et de la croissance peuvent être attribués à l’expression déséquilibrée des gènes liés à l’X entre les sexes, en particulier à certains stades du développement préimplantatoire, quand les 2 chromosomes X peuvent être actifs. Si l’on considère l’organisme entier, les chromosomes sexuels jouent un rôle crucial dans l’établissement du dimorphisme sexuel des fonctions cellulaires. Mais cette expression déséquilibrée n’est pas confinée au développement préimplantatoire, elle intervient aussi ultérieurement, puisque certains gènes portés par le chromosome X échappent à l’inactivation ou sont soumis à empreinte. L’initiation de la différenciation sexuelle dépend du gène SRY, codé par le chromosome Y. Les gènes de détermination du sexe situés sur les chromosomes sexuels peuvent influencer non seulement le développement et le maintien différencié d’organes sexuels secondaires non gonadiques, mais aussi ceux d’organes en dehors du système reproductif, comme le foie, le cerveau [40]. Les hormones ont deux types d’effets : permanents « organisationnels » sur les organes génitaux, le cerveau et peut-être d’autres tissus, et des effets aigus « d’activation », via les hormones ovariennes et testiculaires. L’environnement, sous toutes ses formes, intervient à tous les stades avant et pendant la gestation et après la naissance, via des effets biologiques liés au sexe ou des aspects à proprement parler socioculturels contribuant à sculpter le phénotype social basé sur le genre. Toutes ces composantes envoient des signaux à la machinerie et aux marques épigénétiques, permettant à l’épigénome de chaque cellule de refléter le sexe (le genre) de l’individu (flèches vertes). |
| Dans le texte | |
|
Figure 3. Dimorphisme sexuel des effets intergénérationnels et transgénérationnels. Le premier mode de transmission, de loin le mieux connu et étudié, est somatique. Il résulte de l’influence, sur les cellules somatiques du fœtus et du nouveau-né au cours du développement, de perturbations métaboliques ou nutritionnelles ou d’un stress de la mère pendant la grossesse et l’allaitement. Il peut se reproduire à chaque grossesse et engendrer des effets intergénérationnels. Le second mode de transmission passe par la lignée germinale. Alors que les marques épigénétiques des gamètes s’effacent, au cours de deux phases, la différenciation sexuelle et immédiatement après la fécondation, ces remises à zéro peuvent n’être pas complètes et épargner quelques marques ayant mémorisé le vécu d’un ancêtre. L’altération de certaines marques, non effacées, sur certains gènes dans les cellules germinales, pourrait ainsi être transmise à plusieurs générations successives (F1, F2 via les cellules germinales), sans toutefois que l’on en connaisse les mécanismes. Un dimorphisme sexuel peut dépendre du sexe du parent, de la progéniture et être différent aussi en fonction de la génération (revue dans [4, 41]). |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.