")
")
| Issue |
Med Sci (Paris)
Volume 42, Number 5, Mai 2026
|
|
|---|---|---|
| Page(s) | 421 - 424 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/2026074 | |
| Published online | 29 May 2026 | |
La méthylation de l’ADN détermine la position et la stabilité des centromères humains
DNA methylation shapes human centromere position and stability
Institut Curie, Université Paris Sciences et Lettres (PSL), Sorbonne Université, CNRS UMR 144 et 3664, Paris, France
*
This email address is being protected from spambots. You need JavaScript enabled to view it.
**
This email address is being protected from spambots. You need JavaScript enabled to view it.
L’épigénétique des centromères
La plupart des cellules de notre organisme se divisent tout au long de la vie et, à chaque division cellulaire, le génome est dupliqué. Au-delà de la séquence d’ADN elle-même, les cellules doivent aussi transmettre de multiples couches d’information épigénétique, comme la méthylation de l’ADN, une modification chimique qui influence l’organisation de la chromatine, l’expression des gènes et l’intégrité du génome. Cette méthylation est fréquemment dérégulée dans le cancer, ainsi que dans plusieurs autres maladies [1]. Tous les chromosomes comportent une région unique qui assure la répartition du matériel génétique dupliqué entre les cellules filles lors de la division cellulaire : le centromère [2]. La position du centromère dépend de caractéristiques épigénétiques plutôt que de la séquence d’ADN. Plus précisément, la protéine centromérique A (centromeric protein A, CENP-A), une variante de l’histone H3, marque les centromères et recrute, directement ou indirectement, la plupart des autres protéines centromériques et du kinétochore, c’est à dire l’ensemble des protéines nécessaires pour relier les microtubules et les chromosomes et qui assurent le mouvement de ces derniers durant la méiose et la mitose [3]. Dans l’espèce humaine, CENP-A est déposée sur une séquence d’ADN de 171 paires de bases, appelée α-satellite, qui contient, dans certaines variantes, un court motif de liaison pour la protéine centromérique B (CENP-B) [4]. Les α-satellites sont répétés de nombreuses fois, formant ainsi de longs fragments pouvant atteindre plusieurs millions de paires de bases, et dont la nature répétitive a compliqué l’analyse pendant des décennies. Cependant, les progrès des techniques de séquençage nucléotidique ont permis, en 2022, de déterminer les séquences complètes de l’ADN des centromères, ainsi que certaines de leurs caractéristiques épigénétiques [5]. Ces séquences ont révélé que CENP-A est concentrée dans de petites régions centromériques non méthylées, appelées « régions de creux du centromère » (« centromere dip regions », CDR), qui sont entourées de vastes régions fortement méthylées [6]. Jusqu’à présent, on ignorait si cette disposition de méthylation de l’ADN centromérique jouait un rôle dans le positionnement de CENP-A ou dans la fonction du centromère.
La méthylation de l’ADN centromérique contrôle la liaison et la position des protéines du centromère
Pour comprendre le rôle de la méthylation de l’ADN centromérique, nous avons modifié des cellules humaines afin d’augmenter ou de diminuer cette méthylation spécifiquement aux centromères. Pour diminuer la méthylation centromérique, nous avons fusionné le domaine catalytique de l’enzyme de déméthylation TET1 (ten-eleven translocation 1) avec le domaine de liaison au centromère de CENP-B (CENP-BDBD): l’expression inductible de cette protéine de fusion a provoqué une perte progressive de la méthylation de l’ADN centromérique, proportionnelle au degré d’expression de la protéine. Inversement, l’expression d’une protéine de fusion portant une méthyltransférase bactérienne a permis d’augmenter la méthylation centromérique [7] (Figure 1).
 |
Figure 1 Fonctionnement des systèmes de déméthylation ou d’hyperméthylation expérimentale des centromères. À gauche : le système de déméthylation centromérique est activé dans les cellules par l’induction de l’expression de la protéine de fusion du domaine catalytique de l’enzyme de déméthylation TET1 (ten-eleven translocation 1) avec le domaine de liaison de la protéine CENP-B (centromeric protein B) à l’ADN centromérique. À droite : le système d’hyperméthylation est activé dans les cellules par l’induction de l’expression de la protéine de fusion du domaine catalytique de la méthyltransférase bactérienne MQ1 avec le domaine de liaison à l’ADN de CENP-B. Les deux protéines de fusion possèdent aussi un système de dégradation ciblée, ce qui permet de contrôler leur expression. |
La déméthylation de l’ADN centromérique a augmenté les quantités de protéines CENP-B et CENP-A endogènes aux centromères, tandis que l’hyperméthylation les a réduites. Ces changements ont été constatés à toutes les phases du cycle cellulaire, ce qui indique que la méthylation de l’ADN agit directement sur le recrutement et la liaison des protéines centromériques. Les résultats de tests biochimiques in vitro, en accord avec ceux d’une étude précédente [8], l’ont d’ailleurs confirmé, car nous avons montré que CENP-B se lie plus fortement à son motif d’ADN quand celui-ci n’est pas méthylé. La cartographie des séquences nucléotidiques de l’ADN centromérique sur le génome humain de référence a révélé que les centromères sont presque totalement déméthylés après seulement 4 jours de forte expression de la protéine de fusion CENP-BDBD-TET1, effaçant effectivement les frontières qui définissaient les CDR. Cela a permis à CENP-A de s’étendre au-delà de son domaine habituel en quelques divisions cellulaires. Dans ce court espace de temps, l’accessibilité de la chromatine1 a également augmenté, bien que les marques d’hétérochromatine soient restées en grande partie inchangées. Cela suggère que le rôle principal des motifs de méthylation centromérique, et en particulier celui des frontières des CDR, est de restreindre le positionnement de CENP-A.
La déméthylation du centromère affecte la ségrégation chromosomique et la viabilité cellulaire
Compte tenu du contrôle physiologique de la liaison des protéines centromériques à l’ADN au cours du cycle cellulaire, nous avons évalué les conséquences de la perte de la méthylation de l’ADN au centromère. Une déméthylation rapide (en 2 à 4 jours) a altéré la croissance et la viabilité cellulaires, probablement à cause de l’augmentation des erreurs de ségrégation des chromosomes pendant la mitose que nous avons mise en évidence par imagerie microscopique. Nous avons également constaté une augmentation du nombre des « micronoyaux », corpuscules extranucléaires contenant de l’ADN qui se forment lorsqu’un chromosome ou un fragment chromosomique n’est pas incorporé dans l’un des noyaux des cellules filles après la division cellulaire, et qui sont une source d’instabilité du génome. En effet, grâce au séquençage du génome de cellules uniques, nous avons détecté non seulement des aneuploïdies numériques (nombre anormal de chromosomes), mais également des aneuploïdies structurales, conséquences de cassures d’ADN se traduisant par des translocations non équilibrées, des délétions ou des duplications de grandes régions chromosomiques.
De plus, l’analyse par microscopie d’expansion, une technique d’imagerie de super-résolution [9], a mis en évidence une organisation centromérique anormale et des structures en 3D de CENP-A aberrantes, ce qui est en faveur d’un lien direct entre la perte de méthylation du centromère, la désorganisation et l’extension des régions où CENP-A se lie à l’ADN, et les défauts de ségrégation des chromosomes.
Contrairement à la perte rapide de la méthylation centromérique, lorsque la déméthylation se produit lentement (en 8 à 16 jours), les cellules conservent leur viabilité bien que les quantités de protéines centromériques se stabilisent à des valeurs supérieures aux valeurs normales. Ce constat implique un remodelage progressif de l’architecture centromérique, qui induit une « tolérance » à l’hypométhylation. L’analyse d’un modèle cellulaire du syndrome ICF (immunodéficience, instabilité de l’hétérochromatine centromérique, et anomalies faciales), une maladie caractérisée par une réduction de la méthylation de l’ADN (péri)centromérique [10], a d’ailleurs montré une adaptation similaire sans défaut majeur de la croissance cellulaire.
La méthylation de l’ADN définit donc une « frontière épigénétique » qui détermine la position du centromère et assure le partage équitable des chromosomes mitotiques entre les deux cellules filles (Figure 2). Nous avons montré qu’une déméthylation rapide altère l’architecture centromérique et favorise l’instabilité génomique lors des divisons cellulaires, tandis qu’une déméthylation plus lente permet à la cellule de s’y adapter. Ce constat relie la dynamique de la méthylation de l’ADN à des maladies humaines et souligne son rôle dans la préservation de l’intégrité du génome.
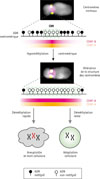 |
Figure 2 Modèle des conséquences moléculaires et cellulaires d’une hypométhylation du centromère. En haut : image d’un chromosome humain témoin avec coloration, par immunofluorescence, des protéines centromériques CENP-A (en jaune) et CENP-B (en magenta). Le schéma représente l’ADN centromérique qui est majoritairement méthylé (boules noires), avec une petite région hypométhylée (« CDR », boules blanches) où se concentre CENP-A. La déméthylation expérimentale des centromères entraîne une altération de leur structure, une augmentation de la quantité des deux protéines, et un élargissement de la région centromérique hypométhylée. Lorsque la déméthylation des centromères se produit rapidement (en 2 à 4 jours), elle entraîne un défaut de ségrégation des chromosomes entre les cellules filles, une aneuploïdie, et la mort cellulaire, tandis qu’une déméthylation plus lente (en 8 à 16 jours) permet au contraire à la cellule de s’adapter à ce nouveau profil épigénétique. |
Perspectives
Nous avons montré que la méthylation régionale différentielle de l’ADN est un déterminant physiologique de l’identité centromérique. En définissant les frontières du dépôt de la protéine CENP-A sur l’ADN et en modulant la liaison de CENP-B, elle préserve la position, et donc la fonction, des centromères. Une perte rapide de la méthylation perturbe cet équilibre et conduit à des erreurs de ségrégation des chromosomes et à l’instabilité du génome lors de la mitose, qui sont deux marqueurs du cancer : en effet, l’hypométhylation des régions d’ADN répétitif est une caractéristique de nombreux types de tumeurs et pourrait contribuer aux réarrangements chromosomiques qui surviennent dans certains cancers. Après cette première étape pour comprendre comment la méthylation de l’ADN et d’autres marques épigénétiques associées, comme les modifications d’histones, contrôlent la fonction du centromère, il conviendra notamment de déterminer comment se forme la région hypométhylée centromérique dite « de creux » (CDR), et de préciser le rôle essentiel de cette modification épigénétique de l’ADN dans la répartition des chromosomes entre les cellules filles.
Liens d’intérêt
Les auteurs déclarent ne pas avoir de lien d’intérêt.
Références
- Greenberg MVC, Bourc’his D. The diverse roles of DNA methylation in mammalian development and disease. Nat Rev Mol Cell Biol 2019 ; 20 : 590-607. [Google Scholar]
- Scelfo A, Fachinetti D. Centromere: A Trojan horse for genome stability. DNA Repair (Amst) 2023 ; 130 : 103569. [Google Scholar]
- McKinley KL, Cheeseman IM. The molecular basis for centromere identity and function. Nat Rev Mol Cell Biol 2016 ; 17 : 16-29. [Google Scholar]
- Masumoto H, Masukata H, Muro Y, et al. A human centromere antigen (CENP-B) interacts with a short specific sequence in alphoid DNA, a human centromeric satellite. J Cell Biol 1989 ; 109 : 1963-73. [Google Scholar]
- Nurk S, Koren S, Rhie A, et al. The complete sequence of a human genome. Science 2022 ; 376 : 44-53. [CrossRef] [PubMed] [Google Scholar]
- Altemose N, Logsdon GA, Bzikadze AV, et al. Complete genomic and epigenetic maps of human centromeres. Science 2022 ; 376 : eabl4178. [Google Scholar]
- Salinas-Luypaert C, Dubocanin D, Lee RJ, et al. DNA methylation influences human centromere positioning and function. Nat Genet 2025 ; 57 : 2509-21. [Google Scholar]
- Tanaka Y, Kurumizaka H, Yokoyama S. CpG methylation of the CENP-B box reduces human CENP-B binding. FEBS J 2005 ; 272 : 282-9. [Google Scholar]
- Sacristan C, Samejima K, Ruiz LA, et al. Vertebrate centromeres in mitosis are functionally bipartite structures stabilized by cohesin. Cell 2024 ; 187 : 3006-23. [Google Scholar]
- Vukic M, Daxinger L. DNA methylation in disease: Immunodeficiency, centromeric instability, facial anomalies syndrome. Essays Biochem 2019 ; 63 : 773-83. [Google Scholar]
L’accessibilité de la chromatine réfère à la capacité contrôlée de protéines ou autres macromolécules présentes dans le noyau à obtenir un accès physique à la chromatine et à l’ADN.
© 2026 médecine/sciences – Inserm
 Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l’utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l’utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Liste des figures
|
Figure 1 Fonctionnement des systèmes de déméthylation ou d’hyperméthylation expérimentale des centromères. À gauche : le système de déméthylation centromérique est activé dans les cellules par l’induction de l’expression de la protéine de fusion du domaine catalytique de l’enzyme de déméthylation TET1 (ten-eleven translocation 1) avec le domaine de liaison de la protéine CENP-B (centromeric protein B) à l’ADN centromérique. À droite : le système d’hyperméthylation est activé dans les cellules par l’induction de l’expression de la protéine de fusion du domaine catalytique de la méthyltransférase bactérienne MQ1 avec le domaine de liaison à l’ADN de CENP-B. Les deux protéines de fusion possèdent aussi un système de dégradation ciblée, ce qui permet de contrôler leur expression. |
| Dans le texte | |
|
Figure 2 Modèle des conséquences moléculaires et cellulaires d’une hypométhylation du centromère. En haut : image d’un chromosome humain témoin avec coloration, par immunofluorescence, des protéines centromériques CENP-A (en jaune) et CENP-B (en magenta). Le schéma représente l’ADN centromérique qui est majoritairement méthylé (boules noires), avec une petite région hypométhylée (« CDR », boules blanches) où se concentre CENP-A. La déméthylation expérimentale des centromères entraîne une altération de leur structure, une augmentation de la quantité des deux protéines, et un élargissement de la région centromérique hypométhylée. Lorsque la déméthylation des centromères se produit rapidement (en 2 à 4 jours), elle entraîne un défaut de ségrégation des chromosomes entre les cellules filles, une aneuploïdie, et la mort cellulaire, tandis qu’une déméthylation plus lente (en 8 à 16 jours) permet au contraire à la cellule de s’adapter à ce nouveau profil épigénétique. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.