")
")
| Issue |
Med Sci (Paris)
Volume 36, Number 1, Janvier 2020
|
|
|---|---|---|
| Page(s) | 12 - 15 | |
| Section | Le Magazine | |
| DOI | https://doi.org/10.1051/medsci/2019253 | |
| Published online | 04 February 2020 | |
Rétablir le métabolisme cérébral du cholestérol est neuro-protecteur dans la maladie de Huntington
Regulation of cholesterol metabolism is neuroprotective in Huntington’s disease
1
Neuroscience Paris Seine, Institut de Biologie Paris-Seine, Paris, France
2
Sorbonne Université, faculté des sciences, Paris, France
3
CNRS UMR8246, Paris, France
4
Inserm U1130, Paris, France
5
Institut du Cerveau et de la Moelle épinière (ICM), Paris, France
6
Grenoble Institut des Neurosciences (GIN), Grenoble, France
* This email address is being protected from spambots. You need JavaScript enabled to view it.
La maladie de Huntington (MH) est une maladie génétique héréditaire de transmission autosomique dominante à pénétrance complète, causée par une expansion anormale du nombre de triplets CAG dans l’exon 1 du gène codant la protéine huntingtine (HTT), dont la séquence d’acides aminés comporte alors un segment poly-glutamine anormalement long dans sa partie N-terminale (HTTm) [1]. La maladie, qui se déclare lorsque le nombre de répétitions CAG est supérieur à 35, entraîne des troubles moteurs, cognitifs, et psychiatriques. Ces symptômes sont liés à la dégénérescence progressive d’une structure du cerveau appelée striatum (composée du noyau caudé et du putamen), qui joue un rôle clé dans la coordination et la planification motrice, et dans l’apprentissage par la récompense [2]. La neuro-dégénérescence s’étend ensuite à d’autres structures, en particulier au cortex cérébral. Il s’agit d’une maladie rare, dont la prévalence est de 3 à 7 cas pour 100 000 individus dans la population caucasienne. En France, elle concerne environ 18 000 personnes, dont 6 000 ont déjà des symptômes de la maladie, et 12 000 sont porteuses du gène muté mais encore asymptomatiques. Bien que la protéine mutée soit exprimée au cours du développement embryonnaire, les premiers signes cliniques apparaissent le plus souvent chez l’adulte entre 30 et 50 ans, mais parfois avant (les formes juvéniles, débutant avant l’âge de 20 ans, représentent 10 % des cas), d’autant plus tôt que le nombre des triplets CAG dans le gène HTT est grand. Les individus à risque peuvent réaliser un test génétique de dépistage, souvent associé à un suivi psychologique. La combinaison des symptômes cognitifs, psychiatriques, et moteurs participe au coût élevé de la prise en charge des individus atteints par cette maladie très invalidante. Les patients doivent en effet faire appel à différents professionnels de santé : médecins généralistes, neurologues, aides à domicile, psychologues, kinésithérapeutes, orthophonistes. Il faut ajouter à cela les coûts de prise en charge sur le long terme. La MH appauvrit les familles et a un impact économique et social important pour la société.
La plupart des traitements actuels de la MH ciblent les symptômes, sans guérir la maladie. Des stratégies plus récentes et en cours d’essais thérapeutiques, adossées à des études chez l’animal, visent à diminuer l’expression du gène HTT, afin de stopper ou ralentir la synthèse de la protéine HTT mutée (HTTm) dans les cellules concernées par la dégénérescence [3]. Les essais en cours incluent des approches ciblant l’expression du gène HTT en utilisant des oligonucléotides « antisens » (antisense oligonucleotides ou ASO, essai de phase III) ou un micro-ARN incorporé dans un vecteur viral (AMT-130, essai de phase I/II) [11] (→).
(→) Voir la Nouvelle de A. Durr, m/s n° 11, novembre 2019, page 834
Ces approches très prometteuses présentent cependant des limites. En effet, elles diminuent à la fois la synthèse de la protéine HTTm, codée par l’allèle muté, mais aussi celle de la protéine HTT normale, dont on sait qu’elle est importante pour le fonctionnement des neurones. Des approches ciblant uniquement l’expression de l’allèle HTT muté sont en cours d’étude : ASO ciblant spécifiquement cet allèle grâce aux polymorphismes nucléotidiques identifiés, techniques d’édition du génome (utilisant des nucléases à doigts de zinc ou la technique CRISPR-Cas9), ou autres approches de thérapie génique.
Malgré ces progrès encourageants non seulement pour la communauté des patients, mais également pour celle des chercheurs et cliniciens, il est essentiel de poursuivre les recherches pour mieux comprendre et mieux traiter cette maladie. Des études par imagerie cérébrale chez des individus porteurs de l’allèle muté montrent que l’atrophie du striatum commence une quinzaine d’années avant l’apparition des premiers symptômes cliniques, et s’étend progressivement à d’autres régions cérébrales [3]. Une intervention thérapeutique durant cette période pré-symptomatique semble donc essentielle en vue d’une neuro-protection. L’utilisation de « biomarqueurs » de la MH devrait permettre de suivre les stades très précoces de la maladie, et d’établir ainsi des fenêtres d’intervention thérapeutique adaptées. Ces biomarqueurs (expression de la protéine HTT mutée et de la protéine des neurofilaments NFL dans le liquide céphalo-rachidien et le sang) sont aussi utilisés pour suivre l’efficacité des essais thérapeutiques [3].
Plusieurs modèles précliniques ont été utilisés pour étudier les processus qui conduisent à la neuro-dégénérescence dans la MH. Il apparaît ainsi que la mutation de la protéine HTT et/ou la perte d’expression de la protéine HTT normale entraîne, précocement, de multiples dysfonctionnements cellulaires incluant des dérégulations transcriptionnelles, des altérations du métabolisme énergétique, de la transmission synaptique et du transport axonal, ainsi qu’un défaut de clairance de la protéine HTTm mal repliée, qui peut alors s’agréger et devenir toxique pour la cellule [4]. Des altérations de l’homéostasie du cholestérol cérébral ont été récemment décrites [5]. Le cholestérol est un composant essentiel de la membrane plasmique (où il contribue notamment à la formation des radeaux lipidiques), de la myéline, et des vésicules synaptiques. Une perturbation de son homéostasie peut donc avoir de graves conséquences sur le fonctionnement neuronal, comme en témoignent les maladies neuro-développementales et neuro-dégénératives, telles que le syndrome de Smith-Lemli-Opitz et la maladie de Niemann-Pick type C, dues à des mutations affectant certains enzymes de son métabolisme [5]. La synthèse et le renouvellement du cholestérol présent dans le système nerveux central sont réalisés in situ car le cholestérol synthétisé par les organes périphériques ne traverse pas la barrière hémato-encéphalique [5] (Figure 1). Dans le cerveau adulte, ce sont principalement les astrocytes qui synthétisent le cholestérol, par une série de réactions enzymatiques à partir de l’acétylCoA. Une apolipoprotéine (ApoE) transporte alors le cholestérol dans les neurones où, en tant que composant des membranes cellulaires, il joue un rôle essentiel dans le fonctionnement des synapses et la transmission du signal nerveux. Ce sont les neurones qui réalisent la première étape de sa dégradation par l’enzyme cholestérol 24-hydroxylase (CYP46A1), qui catalyse la conversion du cholestérol en 24 (S)-hydroxycholestérol (24S-OHC). Ce catabolite passe la barrière hémato-encéphalique et est dégradé dans le foie, mais il agit également dans le cerveau, en se liant aux récepteurs nucléaires de type liver X receptors (LXR), pour contrôler la transcription de gènes importants pour le métabolisme du cholestérol et du gène codant ApoE. Le métabolisme du cholestérol dans le système nerveux central des patients atteints de MH est considérablement altéré : diminution de l’expression des enzymes-clés de la biosynthèse du cholestérol, telles que la 3-hydroxy-3-méthyl-glutaryl-coenzyme A réductase (HMGCR), la lanostérol 14-α déméthylase (CYP51), et la 7-déshydrocholestérol réductase (DHCR7), associée à celle des précurseurs du cholestérol, le lanostérol et le lathostérol [5]. Des observations similaires ont été faites dans des modèles cellulaires de la MH in vitro, mais également dans des modèles murins de la maladie. Cette atteinte globale de la voie de synthèse du cholestérol est associée à une diminution de son catabolisme, puisque la concentration plasmatique de 24S-OHC diminue précocement chez les patients MH, avec une évolution parallèle à celle de l’atrophie du striatum [6]. Ainsi, la concentration plasmatique de ce catabolite du cholestérol pourrait constituer un biomarqueur intéressant pour suivre l’évolution de la maladie à des stades pré-symptomatiques [6].
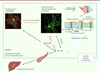 |
Figure 1. Métabolisme du cholestérol dans le cerveau. Le cholestérol est synthétisé par les astrocytes et transporté par l’apolipoprotéine ApoE vers les neurones, dans le fonctionnement desquels sa contribution est majeure. Il est transformé localement par l’enzyme CYP46A1 en 24S-OHC, qui traverse la barrière hémato-encéphalique et gagne le foie, où il est catabolisé. Dans le cerveau, le 24S-OHC se lie aux récepteurs nucléaires LXR au sein des neurones et des astrocytes, régulant ainsi l’expression de gènes importants pour le métabolisme du cholestérol et son transport. |
Dans leur ensemble, ces études suggèrent qu’une restauration du métabolisme du cholestérol pourrait avoir un rôle neuro-protecteur dans la MH. E. Cattaneo et al. ont mis au point des nanoparticules chargées de cholestérol qui, injectées par voie intrapéritonéale chez la souris, permettent de délivrer le cholestérol dans plusieurs régions cérébrales [7]. Ces nanoparticules ont été utilisées dans un modèle murin de la MH, les souris transgéniques R6/2, qui surexpriment un fragment N-terminal de la protéine HTTm, fragment comportant 168 répétitions de glutamine. Chez ces souris, l’administration répétée de nanoparticules chargées de cholestérol, à des stades pré-symptomatiques, améliore l’activité synaptique des neurones du striatum et les fonctions cognitives. Cependant, les fonctions motrices ne sont pas restaurées et les anomalies neurohistopathologiques persistent. En outre, cette stratégie ne permet pas de restaurer le métabolisme global du cholestérol, puisque les taux de son précurseur, le lathostérol, et de 24S-OHC restent diminués. Nous avons alors émis l’hypothèse que la production anormale de 24S-OHC dans la MH était associée à un défaut d’expression et d’activité de CYP46A1. Nous avons montré la baisse du niveau d’expression de CYP46A1 dans des extraits post-mortem de putamen de patients atteints de MH, dans des cellules de lignées striatales exprimant HTTm, ainsi que dans le striatum des souris R6/2 [8]. La restauration de l’expression du gène Cyp46a1 dans des neurones du striatum des souris R6/2, par transfert de l’ADNc correspondant à l’aide d’un vecteur de type adénovirus associé (AAVrh10), réduit le dysfonctionnement neuronal et la toxicité induits par HTTm. Les capacités motrices des souris R6/2 ainsi traitées sont améliorées, les taux de cholestérol et de certains de ses précurseurs sont normalisés, l’atrophie neuronale est moindre, et la quantité d’agrégats de HTTm est diminuée. Dans leur ensemble, ces résultats ont montré qu’une diminution du catabolisme du cholestérol due au déficit en CYP46A1 intervient très précocement dans la pathogénèse de la MH et participe au dysfonctionnement neuronal.
Dans une nouvelle étude, nous avons élucidé les mécanismes qui participent à la neuroprotection produite par la restauration locale de l’expression de CYP46A1 en utilisant un autre modèle murin de la MH, caractérisé par une évolution plus chronique de la maladie : la souris zQ175. Ce modèle, obtenu par une stratégie de knock-in, reproduit plus fidèlement l’anomalie génétique de la maladie humaine, avec la possibilité d’étudier les souris hétérozygotes pour l’allèle muté. Chez ces souris, l’injection du virus AAVrh10 recombinant qui permet de restaurer le taux d’expression de CYP46A1 dans les neurones du striatum a permis de rétablir des fonctions motrices normales. Des études neurohistopathologiques ont confirmé le rôle neuroprotecteur de CYP46A1 en montrant une restauration des marqueurs neuronaux, un rétablissement des arborisations dendritiques, et une clairance des agrégats de HTTm. Une étude transcriptomique à grande échelle par la technique RNAseq indique que la réexpression de CYP46A1 dans les neurones du striatum permet de rétablir l’expression de nombreux gènes impliqués dans la survie neuronale ainsi que dans l’élimination de HTTm. La réexpression de CYP46A1 agit également favorablement sur les fonctions synaptiques et le transport axonal du facteur neurotrophique BDNF (brain-derived neurotrophic factor). Enfin, nous montrons que cette réexpression, en régulant la production de stérols (lanostérol, desmostérol) en amont du cholestérol, stimule l’autophagie et le système ubiquitine-protéasome, deux mécanismes cellulaires importants pour la dégradation des protéines toxiques comme l’est HTTm (Figure 2).
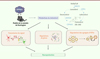 |
Figure 2. La restauration locale de l’expression de CYP46A1 dans le cerveau atténue de multiples dysfonctionnements cellulaires dans un modèle murin de la maladie de Huntington. En régulant le métabolisme du cholestérol, la réexpression de CYP46A1 dans des neurones du striatum à l’aide d’un vecteur viral améliore de nombreuses fonctions cellulaires détériorées par la huntingtine mutée (HTTm), toxique pour les neurones : amélioration de la transmission du signal entre les neurones, rétablissement de l’expression de nombreux gènes, dégradation des agrégats formés par HTTm. La restauration de ces mécanismes cellulaires est à l’origine de l’effet neuroprotecteur et des améliorations comportementales chez la souris modèle de MH. |
Ces résultats [9] mettent en évidence de nouveaux mécanismes neuroprotecteurs liés à l’activité de CYP46A1 et associés à une régulation globale du métabolisme cérébral du cholestérol, avec une compensation de multiples dysfonctionnements cellulaires associés à l’évolution lente de la MH. De nouvelles stratégies thérapeutiques ciblant le métabolisme du cholestérol pourraient ainsi être envisagées dans la MH, mais aussi dans d’autres maladies neurodégénératives pour lesquelles des perturbations de ce métabolisme ont été décrites, en particulier dans la maladie d’Alzheimer. La restauration de l’expression de CYP46A1 s’est d’ailleurs avérée bénéfique dans des modèles murins de cette maladie [10].
Liens d’intérêt
J. Caboche et S. Betuing sont co-inventeurs d’un brevet qui a été licencié à la société Brain Vectis.
R. Kacher déclare n’avoir aucun lien d’intérêt concernant les données publiées dans cet article.
Remerciements
Notre laboratoire est financé par les organismes suivants : centre national pour la recherche scientifique, Inserm, Sorbonne université, faculté des sciences, agence nationale de la recherche (CYPHUNT : convention 13-BXV1-0022-01), association France myopathie-téléthon (référence 20763), European Huntington’s disease network (application number 0867), fondation Groupama « vaincre les maladies rares ».
Références
- The Huntington’s Disease Collaborative Research Group A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington’s disease chromosomes. Cell 1993 ; 72 : 971–983. [CrossRef] [PubMed] [Google Scholar]
- Vonsattel JP Myers R H Stevens TJ et al. Neuropathological classification of Huntington’s disease. J Neuropathol Exp Neurol 1985 ; 44 : 559–577. [CrossRef] [PubMed] [Google Scholar]
- Ross CA Tabrizi SJ Huntington’s disease: from molecular pathogenesis to clinical treatment. Lancet Neurol 2011 ; 10 : 83–98. [CrossRef] [PubMed] [Google Scholar]
- Roze E Saudou F Caboche J Pathophysiology of Huntington’s disease: from huntingtin functions to potential treatments. Curr Opin Neurol 2008 ; 21 : 497–503. [CrossRef] [PubMed] [Google Scholar]
- Karasinska JM Hayden MR Cholesterol metabolism in Huntington disease. Nat Rev Neurol 2011 ; 7 : 561–572. [CrossRef] [PubMed] [Google Scholar]
- Leoni V Mariotti C Tabrizi SJ et al. Plasma 24S-hydroxycholesterol and caudate MRI in pre-manifest and early Huntington’s disease. Brain 2008 ; 131 : 2851–2859. [CrossRef] [PubMed] [Google Scholar]
- Valenza M Chen JY Di Paolo E et al. Cholesterol-loaded nanoparticles ameliorate synaptic and cognitive function in Huntington’s disease in mice. EMBO Mol Med 2005 ; 7 : 1547–1564. [Google Scholar]
- Boussicault L Alves S Lamaziere A et al. CYP46A1, the rate-limiting enzyme for cholesterol degradation, is neuroprotective in Huntington’s disease. Brain 2016 ; 139 : 953–970. [CrossRef] [PubMed] [Google Scholar]
- Kacher R Lamaziere A Heck N et al. CYP4A1 gene therapy deciphers the role of brain cholesterol metabolism in Huntington’s disease. Brain 2019 ; 142 : 2432–2450. [CrossRef] [PubMed] [Google Scholar]
- Hudry E Van Dam D Kulik W et al. Adeno-associated virus gene therapy with cholesterol 24-hydroxylase reduces the amyloid pathology before or after the onset of amyloid plaques in mouse models of Alzheimer’s disease. Mol Ther 2010 ; 18 : 44–53. [CrossRef] [PubMed] [Google Scholar]
- Durr A. Thérapie par ARN anti-sens dans la maladie de Huntington : un immense espoir et beaucoup d’inconnues. Med Sci (Paris) 2019 ; 35 : 834–836. [CrossRef] [EDP Sciences] [PubMed] [Google Scholar]
© 2020 médecine/sciences – Inserm
 Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l'utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l'utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Liste des figures
|
Figure 1. Métabolisme du cholestérol dans le cerveau. Le cholestérol est synthétisé par les astrocytes et transporté par l’apolipoprotéine ApoE vers les neurones, dans le fonctionnement desquels sa contribution est majeure. Il est transformé localement par l’enzyme CYP46A1 en 24S-OHC, qui traverse la barrière hémato-encéphalique et gagne le foie, où il est catabolisé. Dans le cerveau, le 24S-OHC se lie aux récepteurs nucléaires LXR au sein des neurones et des astrocytes, régulant ainsi l’expression de gènes importants pour le métabolisme du cholestérol et son transport. |
| Dans le texte | |
|
Figure 2. La restauration locale de l’expression de CYP46A1 dans le cerveau atténue de multiples dysfonctionnements cellulaires dans un modèle murin de la maladie de Huntington. En régulant le métabolisme du cholestérol, la réexpression de CYP46A1 dans des neurones du striatum à l’aide d’un vecteur viral améliore de nombreuses fonctions cellulaires détériorées par la huntingtine mutée (HTTm), toxique pour les neurones : amélioration de la transmission du signal entre les neurones, rétablissement de l’expression de nombreux gènes, dégradation des agrégats formés par HTTm. La restauration de ces mécanismes cellulaires est à l’origine de l’effet neuroprotecteur et des améliorations comportementales chez la souris modèle de MH. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.