")
")
| Issue |
Med Sci (Paris)
Volume 22, Number 8-9, Août–Septembre 2006
|
|
|---|---|---|
| Page(s) | 688 - 690 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/20062289688 | |
| Published online | 15 août 2006 | |
Origine des vertébrés : la tunique fait-elle le moine ?
Vertebrate origins: does the tunic make the man ?
1
Laboratoire de paléontologie, phylogénie et paléobiologie-CC064, Institut des Sciences de l’évolution UMR 5554/CNRS, Université Montpellier II, Place Eugène Bataillon, 34095 Montpellier Cedex 05 France
2
Canadian Institute for Advanced Research, Département de biochimie, Centre Robert-Cedergren, Université de Montréal, CP 6128, Succursale Centre-ville, Montréal (Québec), H3C 3J7 Canada
3
Département des Sciences de la vie, Université de Liège, B22, Sart Tilman, 4000 Liège, Belgique
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
**
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
***
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Depuis plus d’un siècle, les biologistes débattent de l’origine des vertébrés, groupe animal auquel appartient l’espèce humaine. Au sein des Deutérostomiens (Figure 1), les vertébrés, avec les Céphalochordés et les Urochordés, forment l’embranchement des Chordés (Figure 2A), notamment caractérisés par la présence d’un tube nerveux dorsal appelé chorde. Le plus célèbre représentant des Céphalochordés est l’Amphioxus, qui ressemble superficiellement à une larve de poisson (Figure 1A), tandis que les Urochordés, également connus sous le nom de Tuniciers, sont des organismes marins à distribution cosmopolite.
 |
Figure 1. Quelques représentants de la biodiversité des animaux deutérostomiens partageant le caractère embryonnaire notable que constitue l’ouverture secondaire de la bouche (d’où leur nom) au cours du développement. A. Avec sa silhouette rappelant une larve de poisson, l’Amphioxus (Branchiostoma lanceolatum) a longtemps fait figure de plus proche parent des vertébrés. B. Parmi les Tuniciers, la Cione intestinale (Ciona intestinalis) est en passe de devenir l’un des animaux modèles les mieux caractérisés sur les plans génomique et développemental [1], notamment après le séquençage de son génome achevé en 2002 [2]. C. L’appendiculaire Oikopleura dioica possède un génome dont la taille réduite (estimée entre 51 et 65 mégabases) en fait le plus petit génome de Chordé connu [3]. Son séquençage en cours au Centre national de séquençage d’Evry (http://www.cns.fr/externe/Francais/Projets/Projet_HG/organisme_HG.html) a déjà permis de produire des informations exploitables sur le plan phylogénétique. D, E. En dépit de leur apparente simplicité, les Échinodermes (ici l’oursin Strongylocentrotus purpuratus) et les Hémichordés (ici l’entéropneuste Saccoglossus pusillus) ont probablement eu des ancêtres dont le plan d’organisation était plus complexe (© photos, avec l’aimable autorisation d’Arjan Gittenberg [www.ascidians.com], Daniel Chourrout et Chris Cameron). |
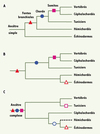 |
Figure 2. Évolution des relations phylogénétiques au sein des Deutérostomiens en fonction du type de caractères utilisés pour les reconstruire. A. Représentation classique reposant sur la morphologie et guidée par l’idée de complexification croissante avec apparition successive des fentes branchiales, de la chorde, puis de la segmentation du mésoderme en somites. B. Image phylogénétique proposée par l’étude de l’ARN ribosomique révélant une proche parenté entre Échinodermes et Hémichordés [5]. Ce schéma évolutif implique que les fentes branchiales étaient vraisemblablement déjà présentes chez l’ancêtre des Deutérostomiens. C. Phylogénie obtenue par l’analyse phylogénomique de 146 gènes nucléaires [4] identifiant les Tuniciers comme les plus proches parents des vertébrés et suggérant une proche parenté entre Céphalochordés et Échinodermes, les Hémichordés n’étant pour l’instant pas inclus dans cette analyse (pointillés). Cette hypothèse suppose que l’ancêtre des Deutérostomiens était probablement bien plus complexe que ce qui a été généralement postulé, avec des fentes branchiales et des caractères de type chordé comme la présence d’une chorde et de somites. L’apparition (symboles pleins) ou la perte (symboles vides) de ces différents caractères au cours de l’évolution est indiquée. |
Avec plus de 2 500 espèces, les Tuniciers présentent une variété morpho-écologique remarquable. La majorité appartient aux Ascidiacés qui, à l’instar de la cione intestinale (Figure 1B), possèdent des larves pélagiques qui se métamorphosent en adultes fixés au substrat. Toutefois, on dénombre également des groupes de Tuniciers exclusivement planctoniques, tels que les Appendiculaires (Figure 1C). Au sein de l’arbre du vivant, les Tuniciers et les Céphalochordés se situent à la frontière entre « invertébrés » et vertébrés. Ils constituent dès lors des modèles de choix pour comprendre l’origine des vertébrés. Outre les Chordés, deux autres embranchements doivent être considérés dans l’étude de cette question. Il s’agit des Échinodermes (lys de mer, ophiures, concombres de mer, oursins et étoiles de mer, Figure 1D) et des Hémichordés (Entéropneustes et Ptérobranches, Figure 1E).
Selon la vision traditionnelle fondée sur la morphologie comparée, l’histoire évolutive des Deutérostomiens est interprétée comme une marche progressive vers la complexité allant des formes « simples » vers les formes plus « complexes », lesquelles « culminent » avec les vertébrés (Figure 2A). Dans ce schéma, les Céphalochordés sont considérés comme les plus proches parents des vertébrés au sein des Chordés, ce qui explique que l’Amphioxus soit largement utilisé comme modèle dans les études d’évolution du développement (Évo-Dévo). Cette proche parenté supposée est cependant essentiellement fondée sur la morphologie externe et sur des caractères partagés, tels que la segmentation du mésoderme en somites, structures embryonnaires situées de part et d’autre du tube neural. Des phylogénies moléculaires fondées sur l’ARN ribosomique [5] ont bousculé cette vision traditionnelle en montrant que les Échinodermes et les Hémichordés sont en fait apparentés (Figure 2B). Cette révélation a logiquement conduit à la réinterprétation de l’évolution de certaines structures morphologiques, telles que les fentes branchiales, que l’on ne retrouve que chez les Hémichordés et les Chordés, mais qui étaient vraisemblablement déjà présentes chez l’ancêtre commun des Deutérostomiens et auraient été perdues chez les Échinodermes. Récemment, tirant parti des séquences génomiques d’Oikopleura dioica représentant un groupe particulier de Tuniciers (les Appendiculaires), nous avons assemblé un grand jeu de données de 146 gènes nucléaires issus de 38 espèces d’animaux et de champignons dans le but de reconstruire les relations de parenté au sein des Deutérostomiens. En identifiant de manière très robuste les Tuniciers comme les plus proches parents des vertébrés dans la biodiversité actuelle, notre étude phylogénomique fait voler en éclats le dogme qui voyait dans les Céphalochordés le groupe-frère des vertébrés (Figure 2C).
Cette découverte inattendue revêt des conséquences majeures pour l’interprétation des données paléontologiques, morphologiques et développementales. En particulier, une proche parenté entre Tuniciers et vertébrés oblige à reconsidérer la présence des somites, classiquement utilisée pour allier Céphalochordés et vertébrés, comme une caractéristique ancestrale des Deutérostomiens qui aurait été réduite secondairement chez les Tuniciers, les Hémichordés et les Échinodermes. Cette hypothèse est en accord avec la possible présence de structures rappelant les myotomes (structures anatomiques issues des somites), chez le plus ancien Tunicier fossile connu [6]. Nos résultats font par ailleurs écho à la récente détection chez les Tuniciers de cellules migratrices de la crête neurale, jusqu’alors considérées comme caractéristiques des vertébrés, alors qu’elles n’ont pour l’instant pas été identifiées chez l’Amphioxus. Dans ce contexte, les Tuniciers apparaissent non plus comme des organismes « primitifs », mais plutôt comme des animaux à la morphologie très différente des autres Chordés car ils se sont adaptés à un mode de vie spécialisé. À l’inverse, les Céphalochordés auraient conservé de nombreux caractères ancestraux.
En rapprochant les Céphalochordés des Échinodermes, nos analyses phylogénomiques suggèrent par ailleurs une hypothèse d’évolution particulièrement intéressante parce qu’elle invalide la monophylie des Chordés. Si cette hypothèse venait à être corroborée par des données supplémentaires, telles que des séquences génomiques d’Hémichordés, elle bouleverserait alors complètement notre vision de l’évolution des Deutérostomiens. En effet, pareille phylogénie impliquerait que l’ancêtre des animaux deutérostomiens ait été un animal beaucoup plus complexe que celui considéré actuellement, avec notamment la présence de fentes branchiales, de somites, d’une chorde et d’un système nerveux dorsal (Figure 2C).
Ces nouveaux résultats devraient donc contribuer au regain d’intérêt dont bénéficient depuis quelques années les Tuniciers. En effet, notre démonstration de la proximité phylogénétique des vertébrés et de ces organismes marins méconnus donne encore plus de sens aux études biomédicales menées sur ces derniers. Ainsi, la récente découverte d’un locus d’histocompatibilité chez l’Ascidie coloniale, Botryllus schlosseri, nous plonge directement aux origines de notre système immunitaire [8] tout en nous permettant de mieux comprendre le fonctionnement des cellules souches [9]. Comptant parmi les organismes producteurs de molécules aux propriétés anti-tumorales [10], les Tuniciers constituent, en outre, un réservoir potentiel de biomolécules naturelles qui ne demande qu’à être exploré.
Remerciements
Cette publication représente la contribution 2006-040 de l’Institut des Sciences de l’évolution de Montpellier (UMR 5554-CNRS). D.B. est chargé de recherche du FNRS.
Références
- Satoh N. The ascidian tadpole larva : comparative molecular development and genomics. Nat Rev Genet 2003; 4 : 285–95. [Google Scholar]
- Dehal P, Satou Y, Campbell RK, et al. The draft genome of Ciona intestinalis : insights into chordate and vertebrate origins. Science 2002; 298 : 2157–67. [Google Scholar]
- Seo HC, Kube M, Edvardsen RB, et al. Miniature genome in the marine chordate Oikopleura dioica. Science 2001; 294 : 2506. [Google Scholar]
- Delsuc F, Brinkmann H, Chourrout D, et al. Tunicates and not cephalochordates are the closest living relative of vertebrates. Nature 2006; 439 : 965–8. [Google Scholar]
- Winchell CJ, Sullivan J, Cameron CB, et al. Evaluating hypotheses of deuterostome phylogeny and chordate evolution with new LSU and SSU ribosomal DNA data. Mol Biol Evol 2002; 19 : 762–76. [Google Scholar]
- Chen JY, Huang DY, Peng QQ, et al. The first tunicate from the early Cambrian of South China. Proc Natl Acad Sci USA 2003; 100 : 8314–8. [Google Scholar]
- Jeffery WR, Strickler AG, Yamamoto Y. Migratory neural crest-like cells form body pigmentation in a urochordate embryo. Nature 2004; 431 : 696–9. [Google Scholar]
- De Tomaso AW, Nyholm SV, Palmeri KJ, et al. Isolation and characterization of a protochordate histocompatibility locus. Nature 2005; 438 : 454–9. [Google Scholar]
- Laird DJ, De Tomaso AW, Weissman IL. Stem cells are units of natural selection in a colonial ascidian. Cell 2005; 123 : 1351–60. [Google Scholar]
- Rinehart KL. Antitumor compounds from tunicates. Med Res Rev 2000; 20 : 1–27. [Google Scholar]
© 2006 médecine/sciences - Inserm / SRMS
Liste des figures
|
Figure 1. Quelques représentants de la biodiversité des animaux deutérostomiens partageant le caractère embryonnaire notable que constitue l’ouverture secondaire de la bouche (d’où leur nom) au cours du développement. A. Avec sa silhouette rappelant une larve de poisson, l’Amphioxus (Branchiostoma lanceolatum) a longtemps fait figure de plus proche parent des vertébrés. B. Parmi les Tuniciers, la Cione intestinale (Ciona intestinalis) est en passe de devenir l’un des animaux modèles les mieux caractérisés sur les plans génomique et développemental [1], notamment après le séquençage de son génome achevé en 2002 [2]. C. L’appendiculaire Oikopleura dioica possède un génome dont la taille réduite (estimée entre 51 et 65 mégabases) en fait le plus petit génome de Chordé connu [3]. Son séquençage en cours au Centre national de séquençage d’Evry (http://www.cns.fr/externe/Francais/Projets/Projet_HG/organisme_HG.html) a déjà permis de produire des informations exploitables sur le plan phylogénétique. D, E. En dépit de leur apparente simplicité, les Échinodermes (ici l’oursin Strongylocentrotus purpuratus) et les Hémichordés (ici l’entéropneuste Saccoglossus pusillus) ont probablement eu des ancêtres dont le plan d’organisation était plus complexe (© photos, avec l’aimable autorisation d’Arjan Gittenberg [www.ascidians.com], Daniel Chourrout et Chris Cameron). |
| Dans le texte | |
|
Figure 2. Évolution des relations phylogénétiques au sein des Deutérostomiens en fonction du type de caractères utilisés pour les reconstruire. A. Représentation classique reposant sur la morphologie et guidée par l’idée de complexification croissante avec apparition successive des fentes branchiales, de la chorde, puis de la segmentation du mésoderme en somites. B. Image phylogénétique proposée par l’étude de l’ARN ribosomique révélant une proche parenté entre Échinodermes et Hémichordés [5]. Ce schéma évolutif implique que les fentes branchiales étaient vraisemblablement déjà présentes chez l’ancêtre des Deutérostomiens. C. Phylogénie obtenue par l’analyse phylogénomique de 146 gènes nucléaires [4] identifiant les Tuniciers comme les plus proches parents des vertébrés et suggérant une proche parenté entre Céphalochordés et Échinodermes, les Hémichordés n’étant pour l’instant pas inclus dans cette analyse (pointillés). Cette hypothèse suppose que l’ancêtre des Deutérostomiens était probablement bien plus complexe que ce qui a été généralement postulé, avec des fentes branchiales et des caractères de type chordé comme la présence d’une chorde et de somites. L’apparition (symboles pleins) ou la perte (symboles vides) de ces différents caractères au cours de l’évolution est indiquée. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.