")
")
| Issue |
Med Sci (Paris)
Volume 42, Number 5, Mai 2026
|
|
|---|---|---|
| Page(s) | 447 - 450 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/2026079 | |
| Published online | 29 May 2026 | |
La taurine entre dans l’arène métabolique de la niche leucémique
Taurine takes the ring in the metabolic arena of the leukemic niche
Université de Toulouse, Centre de recherches en cancérologie de Toulouse, Inserm UMR 1037, CNRS U5077, Toulouse, France
*
This email address is being protected from spambots. You need JavaScript enabled to view it.
Repenser le métabolisme de la niche leucémique à travers la taurine
Depuis plusieurs décennies, la notion de « niche médullaire », soutien des cellules souches hématopoïétiques ou leucémiques, structure notre compréhension de l’hématopoïèse normale et pathologique. Introduite par R. Schofield en 1978, elle a pris une dimension nouvelle au début des années 2000, lorsque les travaux de D. Scadden ont montré que les cellules leucémiques remodèlent leur microenvironnement et transforment la niche médullaire en acteur de la progression tumorale [1]. En effet, la niche médullaire fournit aux cellules souches des signaux essentiels d’adhérence, de survie et de quiescence par des voies de signalisation bien caractérisées, telles que celles impliquant CXCL12 (CXC motif chemokine ligand 12) - CXCR4 (CXC motif receptor 4) [2], SCF (stem cell factor) - récepteur c-KIT [3], ou encore VLA-4 (very late antigen-4) - VCAM-1 (vascular cell adhesion molecule-1) [4]. Au cours de la dernière décennie, cette vision centrée sur la signalisation s’est enrichie d’une dimension métabolique lorsqu’on a découvert que la niche médullaire peut fournir aux cellules souches leucémiques des ressources énergétiques leur permettant de s’adapter aux contraintes métaboliques, notamment sous la pression thérapeutique. Les résultats de plusieurs travaux de recherche ont ainsi montré que les cellules stromales exportent la cystéine afin de maintenir la biosynthèse du glutathion [5], que les adipocytes médullaires libèrent des acides gras utilisés pour l’oxydation mitochondriale dans les cellules leucémiques [6], ou encore que la niche ostéoblastique est remodelée par l’axe kynurénine-HTR1B (serotonin receptor 1B) – SAA (serum amyloid A) – IDO1 (indoleamine 2,3-dioxygenase-1) induit par les cellules leucémiques [7].
Une étude récente [8] apporte un éclairage original sur cette complexité en mettant en évidence l’hétérogénéité non génétique de l’écosystème tumoral ainsi que sa dynamique temporelle. Les auteurs identifient la taurine (acide 2-aminoéthanesulfonique), produite par la niche médullaire au cours du temps, comme un nouveau médiateur clé de ce dialogue métabolique [8]. La taurine est un acide aminé soufré non essentiel présent à l’état libre dans l’organisme, synthétisé à partir de la méthionine et de la cystéine par la cystéine sulfinate décarboxylase (Figure 1). Sa concentration est particulièrement élevée dans l’os, le muscle et le système nerveux, où elle intervient dans le métabolisme, l’homéostasie calcique, et la protection cellulaire. Dans cette étude, la taurine apparaît comme un facteur pro-leucémique, dont la production par la niche médullaire soutient la progression de la maladie.
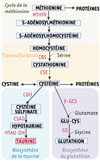 |
Figure 1 Voie de biosynthèse de la taurine. La voie de biosynthèse de la taurine converge avec le cycle de la méthionine, la voie de trans-sulfuration de l’homocystéine et la biosynthèse du glutathion à l’étape de la cystéine. Cette dernière peut également être convertie en cystine ou incorporée dans les protéines. La taurine est synthétisée par l’action séquentielle de trois enzymes : la cystéine dioxygénase (CDO), la cystéine sulfinate décarboxylase (CSAD), et l’hypotaurine déshydrogénase (HTAU-DH). MTHFR : méthylènetétrahydrofolate réductase ; CBS : cystathionine β-synthase ; CSE : cystathionine γ-lyase ; γ-GCS : γ-glutamyl cystéine synthétase ; GS : glutathion synthétase. Figure réalisée avec BioRender. |
L’axe taurine/SLC6A6 dans les leucémies aiguës myéloïdes
Par séquençage nucléotidique de l’ARN à l’échelle de la cellule unique, les chercheurs ont suivi l’évolution des populations stromales non hématopoïétiques de la moelle osseuse au cours de la transition de l’état physiologique vers la leucémie myéloïde naissante, évolutive, et terminale, chez des souris non conditionnées, sans irradiation sublétale préalable à la greffe leucémique, afin de préserver les cellules de la niche médullaire. Afin de construire l’interactome de la niche médullaire leucémique, les données obtenues ont été combinées aux profils transcriptomiques des cellules souches leucémiques humaines. Cette approche intégrative a permis d’identifier les paires récepteur-ligand déterminantes pour le maintien des cellules souches leucémiques au sein du microenvironnement remanié. Les auteurs montrent une expansion des cellules de la lignée ostéoblastique exprimant la cystéine dioxygénase 1 (CDO1), enzyme catalysant la biosynthèse de la taurine, chez les souris leucémiques. Or, l’expression de cette enzyme augmente au cours du temps dans les cellules stromales des patients, lors de la progression d’un syndrome myélodysplasique vers une leucémie aiguë myéloïde secondaire, ainsi qu’entre le diagnostic initial et la rechute d’une leucémie aiguë myéloïde. La délétion conditionnelle de Cdo1 dans les lignées ostéoblastiques murines réduit l’expansion leucémique et prolonge la survie des souris, ce qui établit un lien causal direct entre un métabolite produit par la niche médullaire et la progression de la maladie. Les cellules souches leucémiques sont fortement enrichies en transporteur de la taurine SLC6A6, et l’interaction de la taurine avec son transporteur active la voie RAG (Ras-related GTP binding) – mTOR (mammalian target of rapamycin), ce qui stimule la glycolyse1.
Par des approches génétiques et pharmacologiques, les auteurs montrent que l’axe taurine-SLC6A6 est indispensable à la progression leucémique in vivo, sans effet toxique majeur sur l’hématopoïèse normale. Les résultats de cette étude révèlent également que l’expression de SLC6A6 est plus forte dans les leucémies aiguës myéloïdes résistantes au vénétoclax, et que l’inhibition de SLC6A6 agit en synergie avec cette molécule anti-cancéreuse, positionnant la taurine comme un nouvel agent de vulnérabilité métabolique dans les leucémies résistantes à ce traitement [8]. Il convient d’ajouter que SLC6A6 assure également le transport de la taurine dans la mitochondrie, où elle est impliquée dans les modifications des ARN de transfert nécessaires à la synthèse des protéines mitochondriales et à la prolifération tumorale [9]. Cette localisation mitochondriale de SLC6A6 élargit le champ d’action de la taurine au-delà de sa signalisation cytosolique via mTORC1, et positionne ce transporteur comme un régulateur central du métabolisme énergétique des cellules cancéreuses.
Taurine et plasticité métabolique des cellules souches leucémiques : un nouvel acteur dans la résistance des leucémies aiguës myéloïdes au vénétoclax
Les cellules souches leucémiques utilisent préférentiellement la phosphorylation oxydative (OXPHOS) pour produire l’ATP, et dépendent donc de cette voie métabolique mitochondriale pour leur survie, contrairement aux cellules souches hématopoïétiques et aux cellules leucémiques « matures » (blastes), qui peuvent produire l’ATP par la glycolyse [10]. Cette vulnérabilité des cellules souches leucémiques a orienté les stratégies thérapeutiques vers le ciblage des mitochondries, et est à l’origine du succès des inhibiteurs de la protéine BCL2 (B-cell lymphoma 2), comme le vénétoclax, pour traiter les leucémies aiguës myéloïdes. Pourtant, les échecs thérapeutiques associés à ces médicaments restent fréquents. Des travaux de recherche ont montré que les cellules souches leucémiques qui survivent au traitement par vénétoclax seul ou en association avec l’azacitidine adaptent leur métabolisme énergétique pour maintenir la production d’ATP par OXPHOS, principalement par une augmentation de l’importation d’acides gras grâce au transporteur CD36 (Figure 2) [10-12]. L’identification de la taurine, métabolite produit par la niche médullaire, comme un signal indispensable à l’activation de mTORC1 (mTOR complex 1) et au maintien de la glycolyse dans les cellules souches leucémiques vient compléter ce modèle, en révélant que la dépendance métabolique de ces cellules est finement modulée par leur microenvironnement (Figure 2). D’un côté, les cellules souches leucémiques sont décrites comme pauvres en espèces réactives de l’oxygène, riches en mitochondries fonctionnelles et dépendantes de BCL2, des caractéristiques associées à l’OXPHOS ; de l’autre, l’inhibition expérimentale de l’axe taurine – SLC6A6 – mTORC1 entraîne la diminution des voies glycolytiques ainsi que celle des métabolites associés (comme le pyruvate, le glycéraldéhyde-3-phosphate et le 3-phosphoglycérate), et compromet l’auto-renouvellement de ces cellules, un effet réversible par l’apport exogène de pyruvate ou par une activation constitutive de mTORC1. Ce constat indique que la glycolyse joue un rôle direct dans le maintien et l’adaptabilité des cellules souches leucémiques. Dans ce modèle hybride, l’OXPHOS semble nécessaire à la survie et au contrôle de l’équilibre redox de ces cellules, tandis que des activations transitoires de la glycolyse, orchestrées par la niche médullaire, favoriseraient la compétition clonale et la résistance thérapeutique (Figure 2).
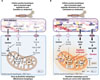 |
Figure 2 Adaptation métabolique des cellules souches leucémiques et de leur microenvironnement dans les leucémies aiguës myéloïdes. (A) Cellule souche leucémique de leucémie aiguë myéloïde de novo sensible au vénétoclax. Ces cellules « naïves » dépendent de la cystéine, des acides aminés qui alimentent le cycle de Krebs, et de la taurine, qui active la voie RAG/mTOR et la glycolyse. Elles expriment BCL2. Leur inflexibilité métabolique les rend vulnérables au vénétoclax, qui induit un effondrement énergétique. (B) Cellule souche leucémique de leucémie aiguë myéloïde réfractaire au vénétoclax ou en rechute. Ces cellules présentent une grande flexibilité métabolique caractérisée par l’importation accrue d’acides gras via le transporteur CD36 et de taurine via le transporteur SLC6A6, induisant respectivement l’oxydation des acides gras et la glycolyse, une reprogrammation métabolique qui confère une résistance au vénétoclax fortement dépendante de la niche médullaire. Qu’elles soient sensibles ou réfractaires au vénétoclax, les cellules souches leucémiques maintiennent une faible concentration des espèces réactives de l’oxygène (phénotype « ROS-low »). GSH : glutathion (forme réduite) ; ATP : adénosine triphosphate ; SIRT3 : sirtuine 3 ; MCL-1 : myeloid cell leukemia-1 ; NAD : nicotinamide adénine dinucléotide ; NAMPT : nicotinamide phosphoribosyl transférase ; MCU : mitochondrial calcium uniporter. Figure réalisée avec BioRender. |
À la différence du glucose ou de la glutamine, la taurine agit comme un métabolite-signal permissif en activant mTORC1 via les GTPases RAG et en induisant l’expression de gènes impliqués dans la glycolyse sous contrôle de facteurs de transcription tels que MYC et via les effecteurs de mTOR. Cette « relecture » du métabolisme des cellules souches leucémiques éclaire le paradoxe selon lequel certaines de ces cellules, bien qu’OXPHOS-dépendantes, développent une résistance au vénétoclax, qui cible pourtant cette vulnérabilité énergétique en agissant sur les mitochondries. L’enrichissement de ces cellules en SLC6A6 suggère que l’activation de la glycolyse dépendante de la niche médullaire pourrait constituer une voie d’échappement métabolique compensant l’inhibition du métabolisme mitochondrial. Si la survie des cellules souches leucémiques repose sur une coopération dynamique entre OXPHOS et glycolyse, cibler une seule de ces deux voies métaboliques est insuffisant pour empêcher cette survie. Cibler l’axe taurine-SLC6A6-mTOR, qui constitue une vulnérabilité « orthogonale » à la vulnérabilité mitochondriale de ces cellules, représente donc une perspective thérapeutique prometteuse, particulièrement dans les leucémies aiguës myéloïdes résistantes au traitement par vénétoclax. Plus généralement, les résultats des travaux de recherche récents plaident pour des stratégies thérapeutiques intégratives qui tiennent compte des flux métaboliques changeants imposés par la niche médullaire, plutôt que des seules propriétés intrinsèques des cellules leucémiques.
Vers un modèle d’écosystème métabolique
La compréhension actuelle du métabolisme des cellules souches leucémiques dépasse donc largement l’opposition réductrice entre OXPHOS et glycolyse, pour rendre compte de la plasticité bioénergétique de ces cellules. Celle-ci repose sur un répertoire métabolique modulable en fonction des contraintes imposées par leur niche médullaire, ainsi que sur l’induction de la fission mitochondriale et de la mitophagie [13]. Un transfert de mitochondries des cellules stromales vers les blastes leucémiques à travers des nanotubes a par ailleurs été montré, contribuant à la reprogrammation métabolique et à la résistance thérapeutique [14]. La reprogrammation du métabolisme des lipides dans certaines cellules souches leucémiques illustre également la richesse de ce répertoire métabolique [15] (→).
(→) Voir m/s n° 12, 2023, page 917
Dans ce contexte, l’identification de la taurine comme ressource des cellules souches leucémiques issue du compartiment ostéoblastique de la moelle osseuse vient élargir le spectre des métabolites fournis par le microenvironnement et qui contribuent à leurs programmes adaptatifs. Les découvertes récentes invitent donc à repenser la résistance thérapeutique des cellules leucémiques, non plus comme une propriété intrinsèque, mais comme l’émergence d’interactions métaboliques dynamiques au sein d’une niche qui s’apparente à un écosystème co-évolutif, structuré dans le temps et dans l’espace.
Perspectives médicales
La taurine est considérée jusqu’à présent comme une molécule thérapeutique dépourvue d’effets indésirables majeurs, et suscite un intérêt croissant pour sa capacité non seulement à prévenir les dommages mitochondriaux, mais aussi à accroître la performance musculaire et la formation osseuse, ainsi qu’à améliorer le fonctionnement cardiaque et hépatique et le métabolisme énergétique du tissu adipeux. De ce fait, elle est largement consommée par le biais de boissons énergisantes et de compléments alimentaires dans le but d’améliorer les performances physiques et cognitives. Les résultats de la recherche sur les cellules souches leucémiques que nous avons présentés soulèvent la question du risque potentiel, chez les personnes atteintes de leucémie, d’une supplémentation en taurine qui pourrait favoriser la progression de la maladie et diminuer l’efficacité de la chimiothérapie. Il importe de noter que l’effet de la taurine semble dépendre du type de cancer. La capacité de la taurine à renforcer la réponse immunitaire anti-tumorale dans le cancer gastrique pourrait justifier une supplémentation alimentaire dans ce contexte [16]. À ce jour, aucune recommandation médicale ne préconise une augmentation ou une restriction de l’apport alimentaire en taurine chez les personnes atteintes de leucémie aiguë myéloïde, en attendant une évaluation rigoureuse de la pertinence médicale des résultats expérimentaux obtenus chez la souris.
La même équipe de chercheurs a montré que l’expression de SLC6A6, et par extension, l’importation de taurine, constitue un déterminant commun à la leucémie aiguë myéloïde, à la leucémie myéloïde chronique et aux syndromes myélodysplasiques, maladies qui impliquent toutes des cellules souches dysfonctionnelles de la moelle osseuse [8]. Ces chercheurs suggèrent que le métabolisme de la taurine représente une vulnérabilité partagée par ces cellules souches, susceptible d’être une cible thérapeutique dans un spectre élargi de néoplasies myéloïdes. Les transporteurs de la taurine comme SLC6A6 sont accessibles pharmacologiquement. Leur inhibition, testée à la fois dans des modèles murins de leucémie aiguë myéloïde et dans des modèles de xénogreffes dérivées de patients atteints de cette leucémie, potentialise l’effet du vénétoclax, tout en épargnant les cellules souches hématopoïétiques normales, suggérant une sélectivité thérapeutique exploitable [8]. Enfin, la combinaison d’inhibiteurs de SLC6A6 avec des agents ciblant l’OXPHOS, l’équilibre redox, ou le métabolisme lipidique pourrait réduire la plasticité métabolique des cellules souches leucémiques et limiter l’émergence de clones résistants au traitement.
Liens d’intérêt
L’auteure déclare ne pas avoir de lien d’intérêt.
Références
- Morrison SJ, Scadden DT. The bone marrow niche for haematopoietic stem cells. Nature 2014 ; 505 : 327-34. [Google Scholar]
- Sugiyama T, Kohara H, Noda M, et al. Maintenance of the hematopoietic stem cell pool by CXCL12-CXCR4 chemokine signaling in bone marrow stromal cell niches. Immunity 2006 ; 25 : 977-88. [Google Scholar]
- Zhang J, Niu C, Ye L, et al. Identification of the haematopoietic stem cell niche and control of the niche size. Nature 2003 ; 425 : 836-41. [Google Scholar]
- Matsunaga T, Takemoto N, Sato T, et al. Interaction between leukemic-cell VLA-4 and stromal fibronectin is a decisive factor for minimal residual disease of acute myelogenous leukemia. Nat Med 2003 ; 9 : 1158-65. [Google Scholar]
- Jones CL, Stevens BM, D’Alessandro A, et al. Inhibition of amino acid metabolism selectively targets human leukemia stem cells. Cancer Cell 2018 ; 34 : 724-40.e4. [Google Scholar]
- Ye H, Adane B, Khan N, et al. Leukemic stem cells evade chemotherapy by metabolic adaptation to an adipose tissue niche. Cell Stem Cell 2016 ; 19 : 23-37. [Google Scholar]
- Galán-Díez M, Borot F, Ali AM, et al. Subversion of serotonin receptor signaling in osteoblasts by kynurenine drives acute myeloid leukemia. Cancer Discov 2022 ; 12 : 1106-27. [Google Scholar]
- Sharma S, Rodems BJ, Baker CD, et al. Taurine from tumour niche drives glycolysis to promote leukaemogenesis. Nature 2025 ; 644 : 263-72. [Google Scholar]
- Li L, You J, Chai ZQ, et al. SLC6A6 imports taurine into mitochondria to sustain mitochondrial translation and tumour growth. Nat Metab 2026 ; doi: 10.1038/s42255-026-01455-6. [Google Scholar]
- Jones CL, Inguva A, Jordan CT. Targeting energy metabolism in cancer stem cells: Progress and challenges in leukemia and solid tumors. Cell Stem Cell 2021 ; 28 : 378-93. [Google Scholar]
- Jones CL, Stevens BM, Pollyea DA, et al. Nicotinamide metabolism mediates resistance to venetoclax in relapsed acute myeloid leukemia stem cells. Cell Stem Cell 2020 ; 27 : 748-64.e4. [Google Scholar]
- Stevens BM, Jones CL, Pollyea DA, et al. Fatty acid metabolism underlies venetoclax resistance in acute myeloid leukemia stem cells. Nat Cancer 2020 ; 1 : 1176-87. [Google Scholar]
- Bosc C, Saland E, Bousard A, et al. Mitochondrial inhibitors circumvent adaptive resistance to venetoclax and cytarabine combination therapy in acute myeloid leukemia. Nat Cancer 2021 ; 2 : 1204-23. [Google Scholar]
- Marlein CR, Zaitseva L, Piddock RE, et al. NADPH oxidase-2 derived superoxide drives mitochondrial transfer from bone marrow stromal cells to leukemic blasts. Blood 2017 ; 130 : 1649-60. [Google Scholar]
- Birsen R, Lauture L, Sarry JE, et al. Ferroptose, métabolisme lipidique, C/EBPα et résistance thérapeutique dans les leucémies aiguës myéloïdes. Med Sci (Paris) 2023 ; 39 : 917-20. [Google Scholar]
- Cao T, Zhang W, Wang Q, et al. Cancer SLC6A6-mediated taurine uptake transactivates immune checkpoint genes and induces exhaustion in CD8+ [Google Scholar]
L’activation de mTORC1 par la voie RAG stimule la glycolyse par une régulation transcriptionnelle, associée à une accumulation des métabolites glycolytiques.
© 2026 médecine/sciences – Inserm
 Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l’utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l’utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Liste des figures
|
Figure 1 Voie de biosynthèse de la taurine. La voie de biosynthèse de la taurine converge avec le cycle de la méthionine, la voie de trans-sulfuration de l’homocystéine et la biosynthèse du glutathion à l’étape de la cystéine. Cette dernière peut également être convertie en cystine ou incorporée dans les protéines. La taurine est synthétisée par l’action séquentielle de trois enzymes : la cystéine dioxygénase (CDO), la cystéine sulfinate décarboxylase (CSAD), et l’hypotaurine déshydrogénase (HTAU-DH). MTHFR : méthylènetétrahydrofolate réductase ; CBS : cystathionine β-synthase ; CSE : cystathionine γ-lyase ; γ-GCS : γ-glutamyl cystéine synthétase ; GS : glutathion synthétase. Figure réalisée avec BioRender. |
| Dans le texte | |
|
Figure 2 Adaptation métabolique des cellules souches leucémiques et de leur microenvironnement dans les leucémies aiguës myéloïdes. (A) Cellule souche leucémique de leucémie aiguë myéloïde de novo sensible au vénétoclax. Ces cellules « naïves » dépendent de la cystéine, des acides aminés qui alimentent le cycle de Krebs, et de la taurine, qui active la voie RAG/mTOR et la glycolyse. Elles expriment BCL2. Leur inflexibilité métabolique les rend vulnérables au vénétoclax, qui induit un effondrement énergétique. (B) Cellule souche leucémique de leucémie aiguë myéloïde réfractaire au vénétoclax ou en rechute. Ces cellules présentent une grande flexibilité métabolique caractérisée par l’importation accrue d’acides gras via le transporteur CD36 et de taurine via le transporteur SLC6A6, induisant respectivement l’oxydation des acides gras et la glycolyse, une reprogrammation métabolique qui confère une résistance au vénétoclax fortement dépendante de la niche médullaire. Qu’elles soient sensibles ou réfractaires au vénétoclax, les cellules souches leucémiques maintiennent une faible concentration des espèces réactives de l’oxygène (phénotype « ROS-low »). GSH : glutathion (forme réduite) ; ATP : adénosine triphosphate ; SIRT3 : sirtuine 3 ; MCL-1 : myeloid cell leukemia-1 ; NAD : nicotinamide adénine dinucléotide ; NAMPT : nicotinamide phosphoribosyl transférase ; MCU : mitochondrial calcium uniporter. Figure réalisée avec BioRender. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.