")
")
| Issue |
Med Sci (Paris)
Volume 42, Number 5, Mai 2026
|
|
|---|---|---|
| Page(s) | 451 - 458 | |
| Section | M/S Revues | |
| DOI | https://doi.org/10.1051/medsci/2026080 | |
| Published online | 29 May 2026 | |
Une nouvelle nomenclature pour les cellules natural killer
A new nomenclature for natural killer cells
1
Aix Marseille Université, CNRS, Inserm, Centre d’immunologie de Marseille-Luminy, Marseille, France
2
APHM, Hôpital de la Timone, Marseille-immunopole profiling platform, Marseille, France
3
Paris-Saclay Cancer Cluster, Villejuif, France
4
Université Paris-Saclay, Gustave Roussy, Inserm, Prédicteurs moléculaires et nouvelles cibles en oncologie, Villejuif, France
5
École Polytechnique, Palaiseau, France
*
This email address is being protected from spambots. You need JavaScript enabled to view it.
**
This email address is being protected from spambots. You need JavaScript enabled to view it.
Résumé
Les récents progrès de la transcriptomique unicellulaire ont profondément renouvelé notre compréhension des cellules immunitaires. Ils ont permis de mieux caractériser l’hétérogénéité des cellules natural killer (NK) et d’établir une classification intégrant l’ensemble de leurs caractéristiques. Ces approches ont aussi permis d’éclairer le comportement des cellules NK infiltrant le microenvironnement tumoral, les mécanismes de leur dysfonctionnement et les limites de leur activité antitumorale. Enfin, elles sont essentielles pour comprendre les mécanismes d’action des nouvelles thérapies ciblant les cellules NK.
Abstract
Recent advances in single-cell transcriptomics have profoundly renewed our understanding of immune cells. They have enabled us to better characterize the heterogeneity of Natural Killer (NK) cells and establish a classification that integrates all their characteristics. These approaches have also shed light on the behavior of NK cells infiltrating the tumor microenvironment, the mechanisms of their dysfunction, and the limits of their antitumor activity. Finally, they are essential for understanding the mechanisms of action of new therapies targeting NK cells.
© 2026 médecine/sciences – Inserm
 Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l’utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l’utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Les cellules natural killer (NK)
Les cellules natural killer (NK) ont été découvertes il y a maintenant 50 ans. Elles ont d’abord été caractérisées par leur capacité d’élimination de cellules tumorales sans l’intervention de cellules présentatrices d’antigène, ni même de la nécessité d’une immunisation préalable [1–3]. Les cellules NK humaines peuvent être identifiées en cytométrie de flux par les marqueurs suivants : CD45+, CD3-, CD14-, CD19-, CD56+ [4]. Les cellules NK se distinguent par leur capacité à reconnaître et à éliminer spontanément les cellules stressées, transformées ou infectées. Elles font partie d’une famille de lymphocytes plus large appelée les cellules lymphoïdes innées (ILC) [5].
Classification conventionnelle des cellules NK
Une dizaine d’années après leur découverte chez la souris, l’être humain et le rat, deux sous-populations majeures de cellules NK humaines ont été caractérisées sur la base de l’expression du marqueur de surface CD56 (cluster de différenciation 56), une isoforme de la molécule d’adhésion cellulaire neuronale N-CAM (neural cell adhesion molecule). Cette classification a permis de distinguer deux sous-populations principales de cellules NK, en fonction du niveau d’expression de ce marqueur : les CD56dim et les CD56bright [6]. D’autres marqueurs de surface permettent d’affiner cette classification, notamment selon leur degré de maturation. Ainsi, les marqueurs de surface CD62L et CD94/NKG2A (NK group 2 family of receptor A) sont associés aux cellules NK précoces. À l’inverse, l’expression des marqueurs CD57 et KIR (Killer-cell immunoglobulin-like receptors) est associée aux cellules NK matures [7–10]. Ce processus de maturation s’accompagne de capacités cytotoxiques accrues [11].
Une population très particulière de cellules NK appelée « cellules NK adaptatives » peut également se multiplier après la reconnaissance spécifique du cytomégalovirus humain (HCMV) par l’intermédiaire de l’engagement du récepteur NKG2C, qui reconnaît notamment un peptide dérivé de la glycoprotéine virale UL40 présentée par le complexe majeur d’histocompatibilité de classe I, HLA-E (human leukocyte antigen - E) [12]. Chez l’être humain, cette réponse déclenche l’expansion oligoclonale d’une population de cellules NK CD94+, NKG2C+ [13]. Ces sous-populations sont caractérisées par une importante reprogrammation épigénétique, se traduisant par une réactivité accrue en cas de réinfection par le HCMV, et par une plus grande longévité [14]. De fait de ces propriétés, ces cellules sont parfois surnommées « cellules NK mémoire » [15] (→).
(→) Voir m/s n° 4, 2013, page 389
Cette population présente également une forte expression des marqueurs CD57, CD2 et des KIR inhibiteurs, qui reconnaissent les molécules du complexe majeur d’histocompatibilité (CMH) de l’individu, ainsi qu’une faible expression de NKG2A, Siglec-7 (sialic acid binding Ig like lectin 7), NKp30 et FcRγ (Fc receptor γ-chain) [13,14,16].
Les marqueurs de résidence sont aussi utilisés pour classer les cellules NK dans les tissus. Ainsi, on peut distinguer des sous-populations NK résidentes sur la base de l’expression à leur surface de CXCR6 (C-X-C motif chemokine receptor 6), CD49a, CD69 ou encore CD103 [17]. Ces molécules favorisent la rétention des cellules NK dans les tissus et empêchent leur retour dans la circulation périphérique, par des mécanismes d’action variés [18]. D’autres sous-populations de cellules NK peuvent être définies sur la base des variations dans le répertoire de leurs récepteurs d’activation, d’inhibition et effecteurs, tels que les récepteurs de la famille SLAM (signaling lymphocyte-activation molecule), NKG2A, NKG2D, les KIR, DNAM-1 (DNAX accessory molecule-1), ainsi que les récepteurs de cytotoxicité naturelle NCR (natural cytotoxicity receptors) comme NKp30 (codé par le gène NCR3), NKp46 (codé par le gène NCR1), qui transmettent des signaux via CD3ζ et FcRγ, et NKp44 (NCR2), qui transmet des signaux via DAP12 (DNAX-activating protein of 12 kDa). Ces variations permettent de discriminer un grand nombre de sous-populations de cellules NK. En résumé, le compartiment des cellules NK comprend un éventail très diversifié de sous-populations. Cette diversité résulte de multiples facteurs, notamment des voies de développement distinctes, une expression variable des KIR, différents stades de différenciation, et l’expansion clonale induite par l’exposition à des agents pathogènes ou des signaux spécifiques aux tissus [19].
La plasticité des cellules NK, qui peut se traduire par des conversions partielles ou totales d’une sous-population de lymphocytes innés (ILC) vers une autre, rend leur classification d’autant plus complexe. En effet, dans des contextes biologiques ou expérimentaux particuliers, tels que l’exposition à des cytokines, les cellules NK peuvent changer profondément, notamment dans la composition des récepteurs présents à leur surface. Ainsi, des expériences menées in vitro et des observations faites in vivo ont montré que, sous l’influence du TGF-β (transforming growth factor β), présent dans de nombreux contextes tumoraux, les cellules NK peuvent acquérir un phénotype de type ILC1. Cette conversion s’accompagne d’une baisse d’expression du facteur de transcription Eomes, et de l’acquisition de marqueurs caractéristiques des ILC1, tels que CD49a, TRAIL ou CD103. Fonctionnellement, cette sous-population présente une activité cytotoxique réduite [20,21].
La cytométrie à haute dimension a joué un rôle déterminant dans l’élucidation des schémas de co-expression des marqueurs de surface dans les populations de cellules NK, le suivi de leurs changements dynamiques au cours du développement et la caractérisation de leurs variations dans les tissus et les conditions pathologiques [22,23]. Cependant, cette approche reste limitée par le nombre de marqueurs pouvant être analysés simultanément, et est intrinsèquement biaisée par le choix du panel d’anticorps. Ces limites soulignent la nécessité de découvrir et d’intégrer des marqueurs supplémentaires afin d’obtenir une caractérisation plus complète de la diversité des cellules NK. L’ensemble de ces marqueurs de surface est présenté dans le Tableau I.
Marqueurs de surface utilisés pour définir les populations de cellules NK humaines dans la classification conventionnelle.
Les cellules NK dans le microenvironnement tumoral
Rôles des cellules NK dans l’immunité antitumorale
Les cellules NK occupent une place bien particulière dans la réponse antitumorale. Tout d’abord, par leur activité cytotoxique directe, qui s’exerce par trois mécanismes complémentaires : 1) la libération de granules lytiques contenant de la perforine, des granzymes et de la granulysine qui induisent principalement l’apoptose (voie intrinsèque) ; 2) l’activation de récepteurs de mort via le ligand de Fas et TRAIL (tumor-necrosis-factor-related apoptosis inducing ligand) qui déclenchent la voie extrinsèque ; et 3) la cytotoxicité dépendante des anticorps via le récepteur CD16 [24]. Après leur activation, les cellules NK contribuent également à l’inflammation, par la production de cytokines telles que l’IFN-γ (interféron-γ), le TNF-α (facteur de nécrose tumorale α), le GM-CSF (facteur de stimulation des colonies de granulocytes et de macrophages) et des chimiokines CCL1, CCL2, CCL3, CCL4, CCL5 et CXCL8, qui peuvent moduler la fonction d’autres cellules immunitaires innées et adaptatives [25]. Elles contribuent ainsi à créer des tumeurs « chaudes » (c’est-à-dire richement infiltrées en cellules immunitaires) en recrutant des cellules dendritiques de type 1 et des cellules T CD8+ [26,27]. La présence des cellules NK dans le microenvironnement tumoral peut également contribuer à éviter l’échappement des tumeurs aux lymphocytes T par la perte du CMH-I. En effet, cette perte de CMH-I est détectée par les cellules NK, ce qui renforce leur capacité à éliminer les cellules tumorales, en raison de l’absence d’activation de leurs récepteurs inhibiteurs normalement sensibles aux molécules de CMH-I. Enfin, les cellules NK contrôlent également les cellules tumorales métastatiques circulantes tout au long de la progression de la maladie [28].
Échappement des cellules tumorales à l’activité des cellules NK
L’activité cytotoxique des cellules NK envers les cellules transformées peut non seulement participer au contrôle de la croissance tumorale, mais également favoriser la sélection de variants tumoraux présentant une résistance. Les tumeurs peuvent alors échapper progressivement aux cellules NK par la modulation des ligands de stress exprimés à leur surface, par exemple en diminuant l’expression des ligands du récepteur activateur NKG2D, tels que MICA/B (major histocompatibility complex-class I chain related proteins A and B). Ces ligands sont exprimés lors de stress cellulaires, tels que les dommages à l’ADN, ou lors d’un stress oxydatif, et leur expression est souvent associée à la transformation des cellules. Elle permet leur reconnaissance et leur élimination par les cellules NK. La répression de l’expression de MICA et MICB est fréquemment observée dans le cas des tumeurs avancées ou métastatiques comparativement aux tumeurs primaires, et elle semble due à une régulation épigénétique [29]. D’autres mécanismes tels que la production par des cellules tumorales de la forme soluble de CD155 qui inhibe la cytotoxicité des cellules NK médiée par DNAM-1 contribuent à l’échappement des tumeurs [30]. D’importantes modifications peuvent également s’opérer directement à la surface des cellules NK en contexte tumoral. Ainsi, la régulation à la baisse de l’expression des récepteurs activateurs tels que NKp30, NKG2D, DNAM-I, NKp46 ou, à l’inverse, la régulation à la hausse de l’expression des récepteurs checkpoint inhibitors (IC), tels que CTLA4 (cytotoxic T-lymphocyte-associated protein 4), TIGIT (T cell immunoreceptor with Ig and ITIM domains), LAG3 (Lymphocyte Activation Gene-3) ou NKG2A, peut entraîner une réduction drastique de l’activité antitumorale des cellules NK à proximité de la tumeur [31,32]. Il a également été montré que le récepteur inhibiteur NKG2A est surexprimé en contexte tumoral, notamment en réponse à la présence d’IL-10, et que ce phénomène est associé à un mauvais pronostic [33]. Le blocage de ce récepteur inhibiteur est aujourd’hui l’objet d’investigations cliniques [34] (→).
(→) Voir m/s n° 12, 2019, page 990
Dysfonctionnement des cellules NK dans le micro-environnement tumoral
L’activité des cellules NK peut être profondément affectée par le microenvironnement tumoral, notamment par une diminution drastique de la production de granzymes, de cytokines et de chimiokines qu’elles sécrètent [24]. Des facteurs solubles tels que le TGF-β, l’adénosine [35], l’activine-A et la PGE-2 (prostaglandine E2) sont connus pour inhiber l’activation et la prolifération des cellules NK [36,37]. Ces facteurs solubles proviennent principalement des cellules tumorales, des fibroblastes associés au cancer, des lymphocytes T régulateurs et de certaines populations myéloïdes présentes dans le microenvironnement tumoral. La cytotoxicité des cellules NK et leur fonction sont également profondément affectées par l’hypoxie du microenvironnement tumoral [38]. Les interactions directes entre les cellules NK et les lymphocytes T régulateurs peuvent également avoir un effet suppressif sur la cytotoxicité des cellules NK [39]. D’autres cellules, telles que les macrophages TREM2+ (triggering receptors expressed on myeloid cells 2) dans le cancer du poumon, peuvent également réduire l’activité des cellules NK en les privant de cytokines stimulantes telles que l’IL-18 et l’IL-15 [40].
Néanmoins, il reste difficile de définir un équivalent au processus bien décrit d’épuisement des lymphocytes T pour les cellules NK. Ce processus résulte d’une stimulation intense et chronique de leur TCR (T-cell Receptor) et se reflète notamment par l’expression des (checkpoint inhibitors)[41]. Pourtant, un modèle de lymphome (RMA-KR1) récemment développé chez la souris a montré que la présence de tumeurs et d’interférons de type I permet également l’expression des inhibiteurs de point de contrôle à la surface des cellules NK, sans que celle-ci soit corrélée à leur dysfonctionnement. Au contraire, les cellules NK exprimant les inhibiteurs de point de contrôle TIM3 (T-cell immunoglobulin and mucin-containing protein-3) et TIGIT (T-cell immunoreceptor with Ig and ITIM domains) sont plus fonctionnelles que celles n’exprimant pas ces récepteurs à leur surface [42]. Ainsi, la compréhension des mécanismes régissant l’activité des cellules NK au sein du microenvironnement tumoral reste encore partielle.
Vers une reclassification des cellules NK : enjeux et bénéfices
L’approche multi-omique combine différentes données, notamment transcriptomiques (ARN transcrits), protéomiques (protéines traduites) et épigénomiques (modifications chimiques de l’ADN et des protéines qui lui sont associées et qui peuvent modifier la structure de la chromatine) désormais accessibles à l’échelle unicellulaire (scRNA-seq, spectrométrie de masse, ATAC-seq). Ces méthodes ont profondément renouvelé l’étude des cellules immunitaires, dont les cellules NK, en révélant leur diversité et de nouvelles sous-populations.
En parallèle des apports de ces techniques de transcriptomique en cellule unique, se sont également développées des nomenclatures hétérogènes, qui rendent les comparaisons difficiles d’une étude à l’autre. Les sous-populations définies dans une étude sont ainsi souvent difficiles à associer, comparer ou mettre en lien avec des sous-populations équivalentes identifiées dans une autre étude. Une classification harmonisée et standardisée est donc nécessaire pour faciliter la compréhension des études ainsi qu’une meilleure communication au sein de la communauté scientifique. Pour élaborer une telle classification et permettre son utilisation par un large public, cette nomenclature doit présenter les caractéristiques résumées dans le Tableau II.
Critères proposés pour l’élaboration d’une classification biologique pertinente.
Proposition d’une nomenclature consensuelle
Dans notre étude [43], nous nous sommes d’abord appuyés sur un jeu de données de CITE-seq2 (cellular indexing of transcriptomes and epitopes) (intégrant donc le transcriptome et les protéines de surface pour chaque cellule) [44] pour caractériser les populations majeures qui composent les cellules NK. Ces données, issues de sang de donneurs sains, nous ont permis de montrer que les cellules NK du sang périphérique étaient divisées en trois populations principales : NK1, NK2 et NK3. Une représentation schématique de cette classification est présentée dans la Figure 1. En intégrant des jeux de données supplémentaires, nous avons pu montrer que la population NK1 contenait 3 sous-populations distinctes, NK1A, NK1B et NK1C. Nous avons également identifié une population dont le profil est intermédiaire entre celui des cellules NK2 et des cellules NK1, que nous avons nommée NKint. Nous avons ensuite utilisé leurs profils transcriptomiques pour décrire les potentialités fonctionnelles de ces cellules. Nous avons ainsi observé que les cellules NK1 présentaient un profil plus cytotoxique et que la nature des cytokines sécrétées différait profondément de celle produite par les cellules NK2 et les cellules NK3. Nous avons également observé que les cellules NK3 du sang comprenaient notamment les cellules NK adaptatives.
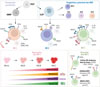 |
Figure 1 Classification des cellules NK. En haut : Les cellules NK1 dérivent des progéniteurs ENKP, tandis que les cellules NK2 proviennent des progéniteurs ILCP. Au milieu : Les cellules NK se répartissent en trois sous-populations principales, chacune définie par des marqueurs de surface (indiqués sur la membrane), des facteurs de transcription (représentés dans le noyau), des granzymes (en bas à droite de chaque cellule) et des cytokines (en haut à droite). En bas à gauche : La trajectoire de maturation des cellules NK1 s’accompagne d’une augmentation progressive de l’activité métabolique, de la cytotoxicité et de la maturation du cytosquelette. En bas à droite : Les cellules NK3 regroupent les cellules NK adaptatives, les cellules NK résidentes tissulaires et certaines cellules NK matures CD57+. ENKP : early NK progenitor, ILCP : innate lymphoid cell progenitor (Figure réalisée avec BioRender). |
Ontologie et trajectoires des cellules NK
Du point de vue de l’ontologie, nos résultats suggèrent que les cellules NK1 et NK2 dérivent de deux progéniteurs distincts. Ces résultats corroborent ceux récemment obtenus chez la souris par Ding et.al [45]. Dans cette étude, les auteurs ont montré que chez la souris, une partie des cellules NK dérivent d’un progéniteur ENKP (early NK progenitor) produisant uniquement des cellules NK, tandis qu’un second progéniteur, ILCP (innate lymphoid cell progenitor), pouvait également générer les autres souspopulations d’ILC (ILC1, ILC2 et ILC3). La conversion des signatures transcriptomiques des cellules NK de souris issues de ces deux progéniteurs en leurs orthologues humains suggère que les NK1 dérivent de l’équivalent humain des ENKP et que les NK2 dérivent de l’équivalent humain des ILCP. Nos données suggèrent donc que chez l’être humain, comme chez la souris, il existe deux progéniteurs de cellules NK distincts qui donnent naissance à deux sous-populations distinctes de cellules NK.
Nos données suggèrent également que les cellules NK1 suivent une trajectoire de maturation en passant successivement par les stades NKint, NK1A, NK1B et enfin NK1C. Cette maturation semble s’accompagner de capacités cytotoxiques accrues, d’une maturation du cytosquelette et d’une activité métabolique accrue, qui permettent de supporter les fonctions effectrices de ces cellules. Pour chacune de ces populations, nous avons également identifié une série de facteurs de transcription et de réseaux de gènes caractéristiques. À l’inverse, les cellules NK2 semblent suivre une trajectoire qui leur est propre et conserver leur identité de cellules NK2. Ces observations sont cohérentes avec l’existence de deux progéniteurs distincts.
Hétérogénéité des cellules NK en situation tumorale
L’approche pan-cancer
La classification NK1, NK2 et NK3 s’applique également aux cellules NK issues des tissus (amygdales, poumon, intestin) et permet d’analyser leur répartition dans le micro-environnement tumoral. En utilisant cette classification sur des données de scRNA-seq provenant de plus de 670 patients couvrant 20 types de cancers différents, issues d’une précédente étude [46], nous avons ainsi observé que le ratio entre les cellules NK1 et les cellules NK2 variait fortement d’un type de cancer à l’autre. Ainsi, nous avons constaté que les cellules NK présentes dans le mélanome sont quasiment exclusivement des cellules NK1 tandis que les neuroblastomes contiennent environ 50 % de cellules NK2. Bien que cette approche offre un bon aperçu de la composition du compartiment cellulaire NK, elle ne tient pas compte des différences de composition entre les tissus sains, ni du dysfonctionnement des cellules NK dans le microenvironnement tumoral. Une étude réalisée avec une méthodologie similaire [47] a permis de répondre partiellement à cette question concernant le dysfonctionnement des cellules NK dans le microenvironnement tumoral, en identifiant une sous-population de cellules NK2 enrichies dans un état stressé dans certains contextes tumoraux. De la même manière, une trajectoire de dysfonctionnement des cellules NK dans le microenvironnement tumoral a été proposée, mais les résultats sont relativement bruités par la présence de nombreux types de cancers et de tissus différents, ce qui rend les comparaisons difficiles. Ainsi, l’approche « pan-cancer », qui combine des données de différents types de cancers et de tissus, est utile pour une vision globale et pour identifier des caractéristiques communes aux cellules NK dans les différentes maladies. En revanche, elle reste limitée lorsqu’il s’agit d’appréhender les spécificités propres à chaque cancer.
L’approche cancer-spécifique
Il convient alors, en particulier lorsqu’on étudie des cellules qui présentent une empreinte tissulaire forte, telles que les cellules NK, d’adopter une approche « cancer-spécifique ». Cela consiste à analyser les cellules dans un contexte tumoral spécifique, en les comparant à des cellules issues d’un tissu sain. On peut ainsi obtenir des données plus homogènes et identifier des signaux plus fins, permettant une meilleure compréhension des mécanismes cellulaires et moléculaires en jeu dans le contexte tumoral.
Le cas du cancer du poumon non à petites cellules
L’approche cancer-spécifique a été appliquée au cancer du poumon non à petites cellules (NSCLC, non-small-cell lung cancer) afin d’analyser la composition du compartiment NK [48]. La classification NK1, NK2 et NK3 décrit l’ensemble des cellules NK présentes dans le microenvironnement tumoral, permettant une résolution fine des sous-populations constitutives. Parmi elles, une fraction de cellules NK3 se caractérise par une forte expression d’ENTPD1 (ectonucleoside triphosphate diphosphohydrolase-1) (CD39), un marqueur associé, chez les lymphocytes T, à des cellules résidentes mémoires réactives à certains antigènes tumoraux [49]. L’analyse transcriptomique indique qu’il s’agit de cellules NK résidentes tissulaires enrichies en contexte tumoral. Ces résultats ont pu être confirmés expérimentalement par un tri en cytométrie de flux suivi d’analyses fonctionnelles. Une fraction de ces cellules exprime l’hétérodimère CD94/NKG2A qui reconnaît les molécules du CMH-1 non classiques HLA-E, et leur activité fonctionnelle est stimulée par le blocage de NKG2A par l’anticorps monoclonal thérapeutique monalizumab [34]. La méthodologie employée pour étudier les cellules NK infiltrées dans les tumeurs du poumon est présentée dans la Figure 2.
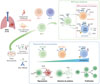 |
Figure 2 Les cellules NK du microenvironnement tumoral dans le NSCLC. En haut : Les cellules NK infiltrées dans le microenvironnement tumoral du NSCLC ont été étudiées en scATACseq. Parmi les cellules NK3, trois sous-populations ont été identifiées. En bas : Les sous-population de cellules NK ont été isolées et traitées avec un mélange de cytokines (IL12, IL15 et IL18) ou avec un anticorps monoclonal anti NKG2A. Les populations NK CD39+ présentent la réponse la plus forte en termes de dégranulation, de production de cytokines et de prolifération. NSCLC : non-small-cell lung cancer (Figure créée avec BioRender). |
Utilisation de la classification pour comprendre les mécanismes de thérapies antitumorales
Les cellules NK possèdent des avantages thérapeutiques distincts de ceux des lymphocytes T, notamment leur capacité à reconnaître les cellules soumises à un stress, leur profil de tolérance favorable lors d’infusions allogéniques, ainsi que leur efficacité potentielle contre les métastases [50,51] (→).
(→) Voir m/s hors série n° 1, 2020, page 50
La classification des cellules NK à l’échelle unicellulaire, couplée à l’analyse transcriptomique, constitue une approche de choix pour disséquer les mécanismes d’action de ces thérapies et en comprendre le fonctionnement. Dans ce contexte, l’analyse en scRNA-seq de cellules NK traitées par un activateur tétraspécifique (CD20-NKCE-IL2v3) a permis de mettre en évidence l’impact de ce dernier sur la fonction et le phénotype des sous-populations NK1, NK2 et NK3. Cet activateur tétra spécifique cible simultanément les récepteurs activateurs NKp46 et CD16 à la surface des cellules NK, le récepteur β de l’IL-2, ainsi que l’antigène tumoral CD20, établissant ainsi un pont physique entre les cellules NK et les cellules tumorales. Ce traitement induit une production soutenue de cytokines, une cytotoxicité accrue et l’émergence de cellules NK de type mémoire, induites par les cytokines [52]. L’analyse a également révélé que la réponse des cellules NK à ce traitement se déroule en deux phases successives : une réponse précoce dominée par les voies de l’interféron, suivie d’une phase proliférative tardive.
Conclusion
Les approches unicellulaires ont profondément renouvelé notre compréhension du compartiment des cellules NK, en révélant une hétérogénéité fonctionnelle et transcriptionnelle d’une ampleur jusque-là insoupçonnée. En permettant l’analyse exhaustive du transcriptome à l’échelle d’une cellule unique, ces techniques intègrent de manière cohérente les dimensions régulatrices, cytotoxiques, cytokiniques et métaboliques des cellules NK, offrant ainsi une cartographie fonctionnelle d’une précision sans précédent. Dans ce contexte, l’établissement d’une nomenclature internationale intégrée constitue une avancée structurante majeure pour le domaine. Elle fournit un cadre commun qui permet d’harmoniser la désignation des sous-populations NK entre les différentes études, de comparer plus rigoureusement les jeux de données générés à l’échelle mondiale et d’en extraire des principes biologiques généralisables.
Par ailleurs, la décomposition fine du compartiment NK en sous-populations distinctes a permis de mieux caractériser l’influence du microenvironnement tumoral sur chacune d’entre elles, ainsi que les effets différenciés des stratégies thérapeutiques qui les ciblent, notamment l’émergence de nouvelles populations fonctionnelles telles que les cellules NK mémoire induites par les cytokines. L’adoption d’une classification harmonisée apparaît ainsi comme une étape indispensable pour structurer durablement l’étude des cellules NK, affiner notre compréhension des interactions immunitaires en contexte tumoral et accélérer le développement de thérapies innovantes. En instaurant un langage commun, cette nomenclature favorise aussi les collaborations internationales et jette les bases d’une immunologie des cellules NK plus intégrative, reproductible et translationnelle.
Liens d’intérêt
Les auteurs déclarent ne pas avoir de liens d’intérêt.
Remerciements
Le laboratoire Éric Vivier du CIML est financé par le Conseil européen de la recherche (CER) dans le cadre du programme Horizon de l’Union européenne (Treatlivmets, convention de subvention n° 101118936), MSDAvenir, Innate Pharma et des subventions institutionnelles accordées au CIML (Inserm, CNRS et Université d’Aix-Marseille). Eric Vivier est titulaire de la chaire « Cellules tueuses naturelles » de la Fondation Gustave Roussy.
Références
- Herberman RB, Nunn ME, Holden HT, et al. Natural cytotoxic reactivity of mouse lymphoid cells against syngeneic and allogeneic tumors. II. Characterization of effector cells. Int J Cancer 1975 ; 16 : 230–9. [Google Scholar]
- Kiessling R, Klein E, Pross H, et al. Natural killer cells in the mouse. II. Cytotoxic cells with specificity for mouse Moloney leukemia cells. Characteristics of the killer cell. Eur J Immunol 1975 ; 5 : 117–21. [Google Scholar]
- Kiessling R, Klein E, Wigzell H. Natural killer cells in the mouse. I. Cytotoxic cells with specificity for mouse Moloney leukemia cells. Specificity and distribution according to genotype. Eur J Immunol 1975 ; 5 : 112–7. [Google Scholar]
- Melsen JE, Themeli M, Ostaijen-Ten Dam MM. Protocol for isolation, stimulation and functional profiling of primary and IPSC-derived human NK cells. Bio Protoc 2020 ; 10 : 3845. [Google Scholar]
- Vivier E, Artis D, Colonna M, et al. Innate lymphoid cells: 10 years on. Cell 2018 ; 174 : 1054–66. [Google Scholar]
- Lanier LL, Le AM, Civin CI, et al. The relationship of CD16 (Leu-11) and Leu-19 (NKH-1) antigen expression on human peripheral blood NK cells and cytotoxic T lymphocytes. J Immunol 1986 ; 136 : 4480–6. [Google Scholar]
- Juelke K, Killig M, Luetke-Eversloh M. CD62L expression identifies a unique subset of polyfunctional CD56dim NK cells. Blood 2010 ; 116 : 1299–307. [Google Scholar]
- Lopez-Vergès S, Milush JM, Pandey S, et al. CD57 defines a functionally distinct population of mature NK cells in the human CD56dimCD16+ NK-cell subset. Blood 2010 ; 116 : 3865–74. [Google Scholar]
- Luetke-Eversloh M, Killig M, Romagnani C. Signatures of human NK cell development and terminal differentiation. Front Immunol 2013 ; 4 : 499. [Google Scholar]
- Yu J, Mao HC, Wei M. CD94 surface density identifies a functional intermediary between the CD56bright and CD56dim human NK-cell subsets. Blood 2010 ; 115 : 274–81. [Google Scholar]
- Bjorkstrom NK, Riese P, Heuts F. Expression patterns of NKG2A, KIR, and CD57 define a process of CD56dim NK-cell differentiation uncoupled from NK-cell education. Blood 2010 ; 116 : 3853–64. [Google Scholar]
- Hammer Q, Ruckert T, Borst EM. Peptide-specific recognition of human cytomegalovirus strains controls adaptive natural killer cells. Nat Immunol 2018 ; 19 : 453–63. [Google Scholar]
- Guma M, Angulo A, Vilches C. Imprint of human cytomegalovirus infection on the NK cell receptor repertoire. Blood 2004 ; 104 : 3664–71. [Google Scholar]
- Ruckert T, Romagnani C. Extrinsic and intrinsic drivers of natural killer cell clonality. Immunol Rev 2024 ; 323 : 80–106. [Google Scholar]
- Narni-Mancinelli E, Ugolini S, Vivier E. Les cellules natural killer: adaptation et mémoire dans le système immunitaire inné. Med Sci (Paris) 2013 ; 29 : 389–95. [Google Scholar]
- Schlums H, Cichocki F, Tesi B. Cytomegalovirus infection drives adaptive epigenetic diversification of NK cells with altered signaling and effector function. Immunity 2015 ; 42 : 443–56. [Google Scholar]
- Sojka DK, Plougastel-Douglas B, Yang L, et al. Tissue-resident natural killer (NK) cells are cell lineages distinct from thymic and conventional splenic NK cells. Elife 2014 ; 3 : e01659. [Google Scholar]
- Bjorkstrom NK, Ljunggren HG, Michaelsson J. Emerging insights into natural killer cells in human peripheral tissues. Nat Rev Immunol 2016 ; 16 : 310–20. [Google Scholar]
- Freud AG, Mundy-Bosse BL, Yu J, et al. The broad spectrum of human natural killer cell diversity. Immunity 2017 ; 47 : 820–33. [Google Scholar]
- Gao Y, Souza-Fonseca-Guimaraes F, Bald T, et al. Tumor immunoevasion by the conversion of effector NK cells into type 1 innate lymphoid cells. Nat Immunol 2017 ; 18 : 1004–15. [Google Scholar]
- Romagnani C, Juelke K, Falco M. CD56brightCD16-killer Ig-like receptor-NK cells display longer telomeres and acquire features of CD56dim NK cells upon activation. J Immunol 2007 ; 178 : 4947–55. [Google Scholar]
- Chretien A-S, Devillier R, Granjeaud S, et al. High-dimensional mass cytometry analysis of NK cell alterations in AML identifies a subgroup with adverse clinical outcome. Proc Natl Acad Sci USA 2021 ; 118 : e2020459118. [Google Scholar]
- Horowitz A, Strauss-Albee DM, Leipold M. Genetic and environmental determinants of human NK cell diversity revealed by mass cytometry. Sci Transl Med 2013 ; 5 : 208ra145. [Google Scholar]
- Coënon L, Geindreau M, Ghiringhelli F, et al. Natural killer cells at the frontline in the fight against cancer. Cell Death Dis 2024 ; 15 : 614. [Google Scholar]
- Paul S, Lal G. The molecular mechanism of natural killer cells function and its importance in cancer immunotherapy. Front Immunol 2017 ; 8 : 1124. [Google Scholar]
- Böttcher JP, Bonavita E, Chakravarty P, et al. NK cells stimulate recruitment of cDC1 into the tumor microenvironment promoting cancer immune control. Cell 2018 ; 172 : 1022–1037.e14. [Google Scholar]
- Allen F, Bobanga ID, Rauhe P, et al. CCL3 augments tumor rejection and enhances CD8+T cell infiltration through NK and CD103+ dendritic cell recruitment via IFNγ. Oncoimmunology 2018 ; 7 : e1393598. [Google Scholar]
- Lopez-Soto A, Gonzalez S, Smyth MJ, et al. Control of metastasis by NK cells. Cancer Cell 2017 ; 32 : 135–54. [Google Scholar]
- Huergo-Zapico L, Acebes-Huerta A, Lopez-Soto A. Molecular bases for the regulation of NKG2D ligands in cancer. Front Immunol 2014 ; 5 : 106. [Google Scholar]
- Okumura G, Iguchi-Manaka A, Murata R, et al. Tumor-derived soluble CD155 inhibits DNAM-1–mediated antitumor activity of natural killer cells. J Exp Med 2020 ; 217 : e20191290. [Google Scholar]
- Merino AM, Kim H, Miller JS, et al. Unraveling exhaustion in adaptive and conventional NK cells. J Leukoc Biol 2020 ; 108 : 1361–8. [Google Scholar]
- Platonova S, Cherfils-Vicini J, Damotte D. Profound coordinated alterations of intratumoral NK cell phenotype and function in lung carcinoma. Cancer Res 2011 ; 71 : 5412–22. [Google Scholar]
- Sun C, Xu J, Huang Q. High NKG2A expression contributes to NK cell exhaustion and predicts a poor prognosis of patients with liver cancer. Oncoimmunology 2017 ; 6 : e1264562. [Google Scholar]
- Cornen S, Andre P, Gauthier L. Les cellules natural killer : des cibles prometteuses dans la thérapie contre le cancer. Med Sci (Paris 2019 ; 35 : 990–2. [Google Scholar]
- Vijayan D, Young A, Teng MWL, et al. Targeting immunosuppressive adenosine in cancer. Nat Rev Cancer 2017 ; 17 : 709–24. [Google Scholar]
- Holt D, Ma X, Kundu N, et al. Prostaglandin E(2) (PGE (2)) suppresses natural killer cell function primarily through the PGE(2) receptor EP4. Cancer Immunol Immunother 2011 ; 60 : 1577–86. [Google Scholar]
- Rautela J, Dagley LF, Oliveira CC. Therapeutic blockade of activin-A improves NK cell function and antitumor immunity. Sci Signal 2019 ; 12 : eaat7527. [Google Scholar]
- Ma S, Caligiuri MA, Yu J. Harnessing natural killer cells for lung cancer therapy. Cancer Res 2023 ; 83 : 3327–39. [Google Scholar]
- Trzonkowski P, Szmit E, Mysliwska J. CD4+CD25+ T regulatory cells inhibit cytotoxic activity of T CD8+ and NK lymphocytes in the direct cell-to-cell interaction. Clin Immunol 2004 ; 112 : 258–67. [Google Scholar]
- Park MD, Reyes-Torres I, LeBerichel J. TREM2 macrophages drive NK cell paucity and dysfunction in lung cancer. Nat Immunol 2023 ; 24 : 792–801. [Google Scholar]
- Wherry EJ, Kurachi M. Molecular and cellular insights into T cell exhaustion. Nat Rev Immunol 2015 ; 15 : 486–99. [Google Scholar]
- Pouxvielh K, Marotel M, Drouillard A, et al. Tumor-induced natural killer cell dysfunction is a rapid and reversible process uncoupled from the expression of immune checkpoints. Sci Adv 2024 ; 10 : eadn0164. [Google Scholar]
- Rebuffet L, Melsen JE, Escalière B, et al. High-dimensional single-cell analysis of human natural killer cell heterogeneity. Nat Immunol 2024 ; 25 : 1474–88. [Google Scholar]
- Hao Y, Hao S, Andersen-Nissen E, et al. Integrated analysis of multimodal single-cell data. Cell 2021 ; 184 : 3573–3587.e29. [Google Scholar]
- Ding Y, Lavaert M, Grassmann S, et al. Distinct developmental pathways generate functionally distinct populations of natural killer cells. Nat Immunol 2024 ; 25 : 1183–92. [Google Scholar]
- Tang F, Li J, Qi L, et al. A pan-cancer single-cell panorama of human natural killer cells. Cell 2023 ; 186 : 4235–4251.e20. [Google Scholar]
- Netskar H, Pfefferle A, Goodridge JP, et al. Pan-cancer profiling of tumor-infiltrating natural killer cells through transcriptional reference mapping. Nat Immunol 2024 ; 25 : 1445–59. [Google Scholar]
- Serger C, Rebuffet L, Sandholzer MT, et al. Integrated single cell analysis identifies CD39+ tumor-associated NK cells with cytotoxic potential in lung cancer. 2025. [Google Scholar]
- Duhen T, Duhen R, Montler R. Co-expression of CD39 and CD103 identifies tumor-reactive CD8 T cells in human solid tumors. Nat Commun 2018 ; 9 : 2724. [Google Scholar]
- Cayatte M, Picant V, Vétizou M, et al. Bringing natural killer cells to the clinic: Opportunities beyond cancer. J Exp Med 2026 ; 223 : e20250612. [Google Scholar]
- Bernard PL, Laletin V, Pastor S, et al. Une piste en immunothérapie du cancer : cibler la signalisation des cellules NK. Med Sci (Paris) 2020 ; 36 : 50–5. [Google Scholar]
- Romee R, Rosario M, Berrien-Elliott MM, et al. Cytokine-induced memory-like natural killer cells exhibit enhanced responses against myeloid leukemia. Sci Transl Med 2016 ; 8 : 357ra123. [Google Scholar]
Lignée lymphomateuse RMA modifiée à l’aide de l’édition génomique CRISPR-Cas9 afin de neutraliser l’expression de B2m. Cette lignée cellulaire n’exprime pas les molécules MHC-I H2Db, H2Kb et B2m à la surface. Les cellules RMA déficientes en B2m ont ensuite été transduites avec un lentivirus exprimant la protéine fluorescente ZsGreen et le ligand NKG2D (Rae1-β) afin d’obtenir la lignée RMA-B2m knockout (KO) Rae1-β+ dénommée RMA-KR (ndlr).
CITE-Seq (indexation cellulaire des transcriptomes et des épitopes) est une méthode basée sur le séquençage qui quantifie simultanément les protéines de surface cellulaire et les données transcriptomiques exprimées dans une seule cellule (ndlr).
Activateur de cellules NK tétraspécifique, il active simultanément les récepteurs d’activation NKp46 et CD16, l’antigène tumoral (CD20) et un récepteur de l’interleukine-2 (via une variante de l’IL-2, IL-2v) (ndlr).
Liste des tableaux
Marqueurs de surface utilisés pour définir les populations de cellules NK humaines dans la classification conventionnelle.
Critères proposés pour l’élaboration d’une classification biologique pertinente.
Liste des figures
|
Figure 1 Classification des cellules NK. En haut : Les cellules NK1 dérivent des progéniteurs ENKP, tandis que les cellules NK2 proviennent des progéniteurs ILCP. Au milieu : Les cellules NK se répartissent en trois sous-populations principales, chacune définie par des marqueurs de surface (indiqués sur la membrane), des facteurs de transcription (représentés dans le noyau), des granzymes (en bas à droite de chaque cellule) et des cytokines (en haut à droite). En bas à gauche : La trajectoire de maturation des cellules NK1 s’accompagne d’une augmentation progressive de l’activité métabolique, de la cytotoxicité et de la maturation du cytosquelette. En bas à droite : Les cellules NK3 regroupent les cellules NK adaptatives, les cellules NK résidentes tissulaires et certaines cellules NK matures CD57+. ENKP : early NK progenitor, ILCP : innate lymphoid cell progenitor (Figure réalisée avec BioRender). |
| Dans le texte | |
|
Figure 2 Les cellules NK du microenvironnement tumoral dans le NSCLC. En haut : Les cellules NK infiltrées dans le microenvironnement tumoral du NSCLC ont été étudiées en scATACseq. Parmi les cellules NK3, trois sous-populations ont été identifiées. En bas : Les sous-population de cellules NK ont été isolées et traitées avec un mélange de cytokines (IL12, IL15 et IL18) ou avec un anticorps monoclonal anti NKG2A. Les populations NK CD39+ présentent la réponse la plus forte en termes de dégranulation, de production de cytokines et de prolifération. NSCLC : non-small-cell lung cancer (Figure créée avec BioRender). |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.