")
")
| Numéro |
Med Sci (Paris)
Volume 31, Numéro 4, Avril 2015
|
|
|---|---|---|
| Page(s) | 383 - 388 | |
| Section | M/S Revues | |
| DOI | https://doi.org/10.1051/medsci/20153104012 | |
| Publié en ligne | 8 mai 2015 | |
Vers une explication des effets mnémoniques des glucocorticoïdes ?
Toward an explanation for the mnemonic effects of glucocorticoids?
Institut de génomique fonctionnelle, département de physiologie, Inserm U661, CNRS UMR 5203, équipe Avenir, 141, rue de la Cardonille, 34070 Montpellier, France
*
freddy.jeanneteau@igf.cnrs.fr
Certains événements sont mieux mémorisés dans un contexte de stress. En effet, les mécanismes engagés par le stress facilitent la formation de la mémoire, mais ils peuvent aussi la détruire. C’est une des difficultés auxquelles se heurtent les stratégies d’identification de cibles médicamenteuses favorisant ou inhibant les effets mnémoniques des glucocorticoïdes. L’imagerie in vivo offre une vision nouvelle des bases structurales de la mémoire pendant les phases d’apprentissage et de la consolidation. La survie des traces de la mémoire dépend de la dose et du rythme d’exposition aux glucocorticoïdes. Ainsi, des rythmes biologiques de glucocorticoïdes anormalement haut ou bas peuvent endommager la plasticité des réseaux neuronaux nécessaires à la formation et à la consolidation de la mémoire. Ces découvertes offrent de nouvelles perspectives de travail pour mieux comprendre le développement de troubles mnésiques et de maladies neuropsychiatriques, telles que la dépression, qui se développent dans un contexte de stress.
Abstract
If the engram of long-term memory is encoded by structural changes of neuronal circuits, they are expected to be present at distant time points after learning, to be specific of circuits activated by learning, and sensitive to behavioral contingencies. In this review we present new concepts that emerged from in vivo imaging studies that tracked the structural bases of the memory trace. A fine balance of spine formation and spine elimination needed for behavioral adaptation to new experience is regulated by glucocorticoids, which are secreted in synchrony with circadian rhythms and in response to stress. Disruption of glucocorticoid oscillations frequently observed in psychiatric disorders like depression and post-traumatic stress produces spine turnover defects and learning disabilities. These new findings provide a new framework for explaining the potent but complex mnemonic effects of glucocorticoids.
© 2015 médecine/sciences – Inserm
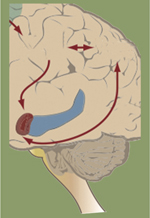
Le cerveau est capable d’une plasticité structurale et fonctionnelle considérable pendant le développement, et cette propriété persiste chez l’adulte [1]. Comme interfaces fonctionnelles entre les neurones, les synapses sont impliquées dans l’assemblage des réseaux neuronaux et l’intégration de l’information. C’est par le contact avec l’axone d’un neurone présynaptique que les épines dendritiques des neurones post-synaptiques filtrent et transforment l’information. Une fois établies, les synapses peuvent persister pendant de très longues périodes. Mais elles peuvent aussi être éliminées en fonction du contexte [2]. L’expérience, par exemple, peut modifier l’organisation des réseaux neuronaux [1, 3]. L’étude du rôle des hormones dans le cerveau a contribué à notre compréhension de ce phénomène important [4, 5]. En particulier, les glucocorticoïdes, dont la sécrétion est sensible au stress, causent un réarrangement réversible des réseaux de connectivité neuronale dans les régions du cerveau impliquées dans l’apprentissage de la récompense, de la cognition, des émotions, de la perception sensorielle [6, 7]. Une hypothèse est que les glucocorticoïdes, bien qu’accessibles à tous les neurones du cerveau, modifieraient les réseaux neuronaux sensibles au contexte environnemental tout en préservant les autres [8, 9]. Rares sont les preuves physiologiques directes d’une telle spécificité d’action. L’imagerie fonctionnelle in vivo, grâce au suivi longitudinal de la dynamique structurale des mêmes neurones au cours des changements de contextes qu’elle permet, valide l’hypothèse d’une coïncidence de détection entre les signalisations induites par les glucocorticoïdes et l’activité neuronale. Reste à déterminer si les traces de la mémoire (engrammes) (voir Glossaire)1 s’établissent par l’intermédiaire de ce type de mécanisme.
Les glucocorticoïdes augmentent le renouvellement des épines dendritiques
Des études classiques de comportement et d’électrophysiologie indiquent que les glucocorticoïdes ont des effets robustes, mais complexes, sur la plasticité synaptique, l’apprentissage et la mémoire [9]. Ces effets sont exprimés sous la forme caractéristique de courbe inversée, car seule une concentration optimale de glucocorticoïde, introduite au bon moment, produit une plasticité maximale. Des concentrations trop faibles ou trop fortes, voire hors contexte, produisent des effets délétères [9–11].
L’imagerie in vivo de la dynamique des épines dendritiques offre une vision complémentaire de celle qu’offre l’électrophysiologie. En effet, une seule injection de glucocorticoïdes permet d’augmenter, en fonction de la dose, la formation et l’élimination des épines dendritiques du cortex sensoriel murin [ . Pour arriver à ce résultat, les glucocorticoïdes empruntent des voies mécanistiques distinctes. Ainsi, la formation des épines est un phénomène rapide (de lx2019;ordre de quelques minutes) alors que l’élimination est un processus beaucoup plus lent (de l’ordre de quelques heures) qui fait intervenir la transcription de nouveaux gènes. Sur la durée, ces phénomènes s’équilibrent, c’est-à-dire qu’il ya autant d’épines formées que d’épines éliminées. Cette forme de plasticité est d’autant plus robuste que l’animal est jeune, mais elle persiste aussi chez l’adulte (Tableau I).
Taux de renouvellement (mesuré sur trois jours) des épines dendritiques dans le cortex murin en réponse à différents changements de l’environnement. Le renouvellement des épines dendritiques augmente lorsque les souris sont maintenues dans un environnement enrichi (large cage + jouets) par rapport a un environnement basal (petite cage standard sans jouets). Les mêmes observations ont été constatées dans le cortex moteur lorsque les souris doivent acquérir des stratégies de coordination motrice pour ne pas chuter d’un Rotarod (3 séances d’apprentissage de 20 essais chacune). Souris adolescentes : âgées de 2 semaines à 1 mois maximum ; souris adultes : âgées de 4 mois au moins.
L’augmentation du renouvellement des épines dendritiques par l’administration de glucocorticoïdes exogènes n’est pas spécifique d’une région du cortex, elle est détectée dans toutes les structures corticales étudiées (les cortex moteur, sensoriel et frontal). Rien d’étonnant puisque les récepteurs des glucocorticoïdes de type GR2 sont ubiquitaires [8, 31]. Au contraire, les récepteurs de type MR, aussi impliqués dans le processus d’élimination des épines, sont peu exprimés dans le cortex. La forte affinité des récepteurs MR pour les glucocorticoïdes, comparée à la faible affinité des récepteurs GR, pourrait compenser cette différence d’expression sans remettre en question un modèle à deux récepteurs
S’ils sont présents en excès, les glucocorticoïdes causent une perte nette d’épines, non seulement des épines nouvelles, mais aussi des épines anciennes, formées au cours du développement. Au contraire, un déficit de glucocorticoïdes, par exemple secondaire à une surrénalectomie, fige la dynamique des épines dendritiques, empêchant le renouvellement de celles-ci (Tableau I). Se pose donc la question de l’utilité physiologique de cette plasticité structurale apparemment non spécifique. L’idée est que l’augmentation de la dynamique des épines dendritiques offre une fenêtre de plasticité pour le réarrangement des réseaux de neurones. En principe, seuls les réseaux neuronaux recrutés par le contexte seraient capables d’exploiter cette fenêtre de plasticité. Des changements d’environnement (par exemple lors d’enrichissement, de stress, d’apprentissage, etc.) pourraient se traduire en réarrangements des réseaux neuronaux s’il y a coïncidence de détection entre l’activité des glucocorticoïdes et l’activité neuronale.
Les bases structurales de la mémoire
C’est pour explorer cette hypothèse de coïncidence que nous avons récemment évalué [13] le rôle des oscillations physiologiques circadiennes des taux de glucocorticoïdes sur la dynamique des épines dendritiques dans plusieurs régions du cortex au cours de l’acquisition et de la consolidation d’une tâche motrice. L’apprentissage moteur n’a pas d’effet sur la plasticité dendritique dans le cortex sensoriel et le cortex frontal. En revanche, dans le cortex moteur, les pics circadiens des taux de glucocorticoïdes facilitent le réarrangement des épines après un apprentissage. Les creux de ces taux circadiens sont, eux, nécessaires à la stabilisation des nouvelles épines qui sont importantes pour la mémoire. Ce réarrangement des épines dendritiques est durable, puisqu’une proportion significative des nouvelles épines formées pendant l’apprentissage survit à long terme. Enfin, la persistance de ces épines est proportionnelle à la performance comportementale évaluée longtemps après l’apprentissage [13]. En principe, c’est parce qu’elles sont stabilisées par l’apprentissage que les épines dendritiques constituent ce que l’on pourrait désigner comme une « unité de stockage de l’information » [14].
Si les traces anatomiques de la mémoire sont codées par des réarrangements entre neurones, ceux-ci devraient, a priori, persister longtemps après l’apprentissage, être spécifiques des neurones activés par l’apprentissage et sensibles au type de tâche apprise. La coïncidence du pic de glucocorticoïdes et de l’apprentissage façonne la connectivité des neurones spécifiquement activés par la tâche comportementale. En revanche, l’absence de coïncidence entre le pic de glucocorticoïdes et l’apprentissage est sans effet (Figure 1).
 |
Figure 1. Le pic circadien de glucocorticoïdes facilite l’apprentissage alors que le creux circadien est nécessaire à la consolidation de la mémoire. Cette phase critique de consolidation dure approximativement une semaine, pendant laquelle toute perturbation des oscillations de glucocorticoïdes a des conséquences néfastes sur la mémoire. En pointillé est illustré l’effet d’une injection de glucocorticoïdes exogènes sur les oscillations des taux de glucocorticoïdes endogènes. Pour limiter les effets du stress, les souris ont préalablement été habituées trois jours de suite sur un rotaRod non motorisé à la procédure qui sera utilisée lors de l’induction du pic de glucocorticoïdes. C’est l’accélération du rotaRod pendant 20 essais/jour à la même heure qui permet aux souris d’apprendre des stratégies de coordination et d’équilibre pour ne pas chuter. L’imagerie in vivo des neurones du cortex moteur lors du rappel de cette tâche après la phase de consolidation de la mémoire indique que la performance comportementale est proportionnelle à la survie des épines dendritiques formées au moment de l’apprentissage et à l’élimination d’épines formées avant l’apprentissage. Contrairement à l’apprentissage, qui est sensible au pic de glucocorticoïdes, la performance lors du rappel ne l’est pas. |
Pendant la semaine consécutive à l’apprentissage, une dérégulation des oscillations des taux de glucocorticoïdes empêche la consolidation de la mémoire et la survie des épines nouvelles formées au moment de l’apprentissage. Ces nouvelles données associent donc les processus de formation des épines dendritiques à l’apprentissage, tandis que la survie et l’élimination des épines dendritiques serviraient à la consolidation et à la rétention de la mémoire. Elles confirment les précédentes observations collectées par des méthodes comportementales et électrophysiologiques [8, 9]. D’ailleurs la rapidité du processus de la formation des épines, comparée à la lenteur des mécanismes de leur survie et de leur élimination, explique pourquoi des variations de l’activité des glucocorticoïdes pendant ces périodes critiques facilitent ou détruisent la mémoire [8]. En effet, la survie des nouvelles épines dendritiques et l’élimination d’épines préexistantes dans le cortex moteur sont deux processus essentiels à l’acquisition et la consolidation d’une nouvelle tâche motrice. Si un ensemble d’épines est façonné au cours d’un apprentissage, la rétention de cet apprentissage devrait dépendre de la survie de cet ensemble d’épines.
Ce modèle a été testé aux limites de la physiologie en évaluant, par exemple, les effets d’une absence ou d’un excès chronique de glucocorticoïdes sur la dynamique des épines dans le cortex moteur pendant l’acquisition et la consolidation d’une tâche motrice [13]. Tout d’abord, une exposition prolongée à des glucocorticoïdes après une tâche d’apprentissage perturbe la rétention de la mémoire et cause une perte des épines formées pendant, et même avant, l’apprentissage. Ainsi, la rétention de la mémoire serait perturbée, car trop de glucocorticoïdes empêcherait la formation de nouvelles épines et exagèrerait l’élimination d’épines préexistantes. Cette situation rappelle les déficits anatomiques, cognitifs et physiologiques détectés chez les patients atteints de dépression majeure [15, 16]. Un déficit en glucocorticoïdes prévient, non seulement la formation, mais aussi l’élimination, d’épines dendritiques ; ce défaut de plasticité structurale se traduirait par un déficit d’apprentissage (Figure 2). Cette situation rappelle les symptômes de patients souffrant de stress post-traumatique [17].
 |
Figure 2. Conséquences d’une perturbation des rythmes circadiens de la sécrétion de glucocorticoïdes sur l’apprentissage et les bases structurales de la mémoire. L’apprentissage au moment du pic de glucocorticoïdes facilite l’acquisition et la consolidation de la mémoire et est corrélé à la formation d’un motif d’épines qui pourrait coder les bases structurales de l’engramme. L’absence chronique des pics de glucocorticoïdes empêche l’apprentissage et la consolidation de la mémoire qui est corrélé à un déficit des processus de formation et d’élimination des épines dendritiques. Au contraire, l’absence de creux circadiens due à un excès chronique de glucocorticoïdes produit des carences cognitives qui sont corrélées avec l’élimination accrue d’anciennes épines dendritiques et un taux de formation quasiment nul. |
Quelques règles de la plasticité d’un engramme cortical induite par des glucocorticoïdes
L’étude des effets mnémoniques des glucocorticoïdes sur la plasticité des épines dendritiques dans un contexte d’apprentissage a donc permis d’établir des règles de la plasticité de l’engramme :
(1) La formation et l’élimination des épines sont des processus dissociés dans le temps, mais équilibrés pour atténuer l’impact de l’apprentissage sur le nombre net d’épines.
(2) La survie des nouvelles épines est corrélée à l’élimination d’épines préexistantes.
(3) La capacité mnésique est proportionnelle à la survie d’un ensemble d’épines.
On observe une exception à ces règles lorsque les rythmes circadiens de la sécrétion de glucocorticoïdes sont perturbés. C’est le cas dans de nombreuses maladies psychiatriques et neurodégénératives qui agissent comme des facteurs aggravants des troubles cognitifs [17–20].
Des mécanismes rapides facilitent la formation des épines dendritiques associées à l’apprentissage
Pour produire cette plasticité des épines dendritiques, les glucocorticoïdes utilisent des mécanismes génomiques et non génomiques qui coïncident avec l’activité neuronale contrôlée par les neurotransmetteurs excitateurs, des facteurs neurotrophiques et des neuropeptides [21, 22]. Des efforts considérables ont été dévolus à la compréhension des mécanismes de plasticité morphologique dans les contextes d’apprentissage et de rétention [23]. Ainsi, la plasticité structurale à court terme dépendrait de modifications post-traductionnelles de protéines associées au cytosquelette, alors que des changements persistants dépendraient également de processus transcriptionnels [24, 25]. Si l’application locale in vivo de glucocorticoïdes par le biais d’une craniotomie augmente rapidement (< 20 minutes) la formation d’épines dendritiques, celle d’antagonistes du récepteur de glucocorticoïdes (GR) empêche cette formation. En revanche, la forte augmentation de la formation d’épines induite par une seule injection de glucocorticoïdes persiste en présence d’actinomycine D, un inhibiteur de la transcription, et après traitement avec un glucocorticoïde synthétique (Cort:BSA) qui ne peut pas accéder aux récepteurs cytoplasmiques3 (voire synaptiques). On peut donc en conclure que le pic de glucocorticoïdes facilite la formation d’épines par un mécanisme rapide non génomique qui dépend de GR (Figure 3).
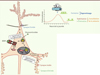 |
Figure 3. Interférence des voies de signalisation des glucocorticoïdes et du BDNF via la phosphorylation des GR. Les glucocorticoïdes utilisent des mécanismes lents et rapides dépendants du GR pour augmenter le renouvellement d’épines dendritiques. La signalisation rapide des GR influence le remodelage du cytosquelette d’actine par la voie LIMK1-cofiline, alors que la signalisation lente des GR phosphorylés cible la transcription de certains gènes, dont celui qui code pour MKP1, une phosphatase influençant le remodelage du cytosquelette de microtubules. Ce mécanisme de signalisation est complexe et dépend du creux circadien du taux de glucocorticoïdes, et des récepteurs GR et MR. Il en résulterait un engramme permettant l’adaptation physiologique de ces réseaux neuronaux aux variations contextuelles. |
Les effets rapides et promnésiques des glucocorticoïdes et la formation de nouvelles épines dépendent de la polymérisation du cytosquelette d’actine par la voie de signalisation LIMK1/cofiline [13]. Le gène LIMK1 (lim domain kinase 1), qui est situé sur le locus q11.23 du chromosome 7 chez l’homme est supprimé, entre autres, par une microdélétion qui touche cette région du chromosome 7 dans le syndrome neurodéveloppemental de Williams, caractérisé par des déficits sévères de plasticité synaptique et des capacités cognitives. Plus spécifiquement, l’invalidation génétique du gène limk1 chez la souris confirme, en partie, la symptomatologie du syndrome de Williams ; elle empêche notamment l’augmentation du taux de renouvellement des épines dendritiques induite normalement par une application de glucocorticoïdes directement sur le cortex par craniotomie [13].
Des mécanismes lents qui facilitent l’élimination et la survie des épines dendritiques associées à l’apprentissage
Les glucocorticoïdes agissent aussi au niveau génomique pour établir le lien entre plasticité synaptique et apprentissage et exercer leurs effets mnémoniques [8]. Puisque seule une petite fraction des épines dendritiques formées pendant l’apprentissage survit au détriment des épines formées avant l’apprentissage, se pose la question de savoir si le choix des épines à éliminer et des épines à maintenir est le résultat d’un mécanisme stochastique ou prédéterminé. Grâce à une approche de criblage génétique, nous avons déterminé que le niveau de BDNF (brain derived neurotrophic factor) influence dramatiquement l’expression du répertoire de gènes sensibles aux glucocorticoïdes, en favorisant le recrutement de cofacteurs spécifiques du récepteur de type GR [26]. Pour cibler certains gènes du répertoire sensibles au récepteur nucléaire, la signalisation du BDNF induit rapidement (de l’ordre de la minute) la phosphorylation du GR sur des sites spécifiques de la liaison de GR à des cofacteurs comme CREB1 (CAMP responsive element binding protein 1) [26] (Figure 3).
Parmi les protéines intermédiaires mises en jeu par la convergence des voies de signalisation du BDNF et des glucocorticoïdes, la phosphatase MKP-1 (mitogen-activated protein [MAP] kinase phosphatase-1), qui désactive la voie de MAP-kinase, facilite le réarrangement structural des neurones. La persistance d’une expression trop élevée du gène correspondant peut, chez l’homme comme chez la souris, conduire à des troubles du comportement cognitif [27]. Au contraire, l’ablation du gène mkp-1 empêche le réarrangement structural des neurones et protège des effets délétères du stress sur l’apprentissage [28, 29]. En outre, manipuler l’expression d’un seul des gènes cibles impliqués dans ces deux voies est suffisant pour induire - ou empêcher - des dysfonctionnements synaptiques engendrés par un déséquilibre des rythmes de sécrétion des glucocorticoïdes.
Conclusions et perspectives
Performance comportementale et réarrangement des épines dendritiques sont des processus optimaux lorsqu’ils coïncident avec le pic du taux de glucocorticoïdes dans le milieu hormonal, et quand les creux oscillatoires sont respectés. Ainsi, la mémoire pourrait être codée par des ensembles d’épines qui résultent des processus de formation et d’élimination. À la suite d’un stress sévère, le nombre d’épines éliminées est supérieur au nombre d’épines formées, le résultat pouvant conduire à des déficits cognitifs, des maladies psychiatriques et à un vieillissement prématuré. En effet, les perturbations du rythme circadien de la sécrétion de glucocorticoïdes dans le milieu hormonal sont très fréquentes dans les maladies psychiatriques, et s’aggravent au cours du vieillissement [27, 30].
Si le concept est clair, il reste cependant de nombreuses questions à élucider. Par exemple, comment une seule hormone, conservée, peut-elle produire des effets aussi complexes, spécifiques et sensibles au développement ? Est-ce qu’empêcher la dynamique des épines dendritiques suffit à protéger les réseaux neuronaux des effets nocifs du stress chronique ? Si c’est le cas, est-ce au détriment de l’apprentissage ? Enfin, la phosphorylation du récepteur des glucocorticoïdes par le BDNF peut-elle représenter un outil mnémonique qui serait exploitable pour lutter contre les effets délétères du stress ?
Liens d’intérêt
L’auteur déclare n’avoir aucun lien d’intérêt concernant les données publiées dans cet article.
Glossaire
Glossaire
Glucocorticoïde : : hormone (cortisol chez l’homme et corticostérone chez la souris ) sécrétée en réponse au stress, à un nouvel environnement (nouveauté) et aux rythmes circadiens.
Stress : : conditions environnementales qui nécessitent une adaptation physiologique de l’organisme. Le stress est intimement lié à l’apprentissage.
Engramme : : traces de la mémoire (anatomiques ou fonctionnelles).
Épine dendritique : : compartiment post-synaptique dynamique composé du cytosquelette d’actine et de récepteurs et d’effecteurs des neurotransmetteurs.
Code d’épines : : le résultat des processus de formation, de survie et d’élimination des épines dendritiques.
GR : : glucocorticoid receptor ; récepteur ubiquitaire de basse affinité pour les glucocorticoïdes. Récepteur nucléaire. Facteur de transcription aussi capable de signalisation synaptique rapide.
MR : : mineralocorticoid receptor ; récepteur limbique de haute affinité pour les glucocorticoïdes. Facteur de transcription aussi capable de signalisation synaptique rapide.
BDNF : : brain-derived neurotrophic factor ; neurotrophine sécrétée de façon dépendante de l’activité neuronale.
Coïncidence de détection : : deux signaux se manifestant ensemble créent une réponse distincte de la somme des deux réponses individuelles
Phosphorylation : : modification protéique post-traductionnelle. La phosphorylation de GR interprète la coïncidence des voies du BDNF et des glucocorticoïdes.
Références
- Yang G, Pan F, Gan WB. Stably maintained dendritic spines are associated with lifelong memories. Nature 2009 ; 462 : 920–924. [CrossRef] [PubMed] [Google Scholar]
- Walsh MK, Lichtman JW. In vivo time-lapse imaging of synaptic takeover associated with naturally occurring synapse elimination. Neuron 2003 ; 37 : 67–73. [CrossRef] [PubMed] [Google Scholar]
- Xu T, Yu X, Perlik AJ, et al. Rapid formation and selective stabilization of synapses for enduring motor memories. Nature 2009 ; 462 : 915–919. [CrossRef] [PubMed] [Google Scholar]
- Maheu FS, Lupien SJ. La mémoire aux prises avec les émotions et le stress : un impact nécessairement dommageable ? Med Sci (Paris) 2003 ; 19 : 118–124. [CrossRef] [EDP Sciences] [PubMed] [Google Scholar]
- De Kloet ER, Joels M, Holsboer F. Stress and the brain: from adaptation to disease. Nat Rev Neurosci 2005 ; 6 : 463–475. [CrossRef] [PubMed] [Google Scholar]
- Moisan MP, Le Moal M. Le stress dans tous ses états. Med Sci (Paris) 2012 ; 28 : 612–617. [CrossRef] [EDP Sciences] [PubMed] [Google Scholar]
- McEwen BS, Morrison JH. The brain on stress: vulnerability and plasticity of the prefrontal cortex over the life course. Neuron 2013 ; 79 : 16–29. [CrossRef] [PubMed] [Google Scholar]
- De Kloet ER, Karst H, Joels M. Corticosteroid hormones in the central stress response: quick-and-slow. Front Neuroendocrinol 2008 ; 29 : 268–272. [CrossRef] [PubMed] [Google Scholar]
- De Quervain DJ, Aerni A, Schelling G, Roozendaal B. Glucocorticoids and the regulation of memory in health and disease. Front Neuroendocrinol 2009 ; 30 : 358–370. [CrossRef] [PubMed] [Google Scholar]
- Roozendaal B, McEwen BS, Chattarji S. Stress, memory and the amygdala. Nat Rev Neurosci 2009 ; 10 : 423–433. [CrossRef] [PubMed] [Google Scholar]
- Joels M, Sarabdjitsingh RA, Karst H. Unraveling the time domains of corticosteroid hormone influences on brain activity: rapid, slow, and chronic modes. Pharmacol Rev 2012 ; 64 : 901–938. [CrossRef] [PubMed] [Google Scholar]
- Liston C, Gan WB. Glucocorticoids are critical regulators of dendritic spine development and plasticity in vivo. Proc Natl Acad Sci USA 2011 ; 108 : 16074–16079. [CrossRef] [Google Scholar]
- Liston C, Cichon JM, Jeanneteau F, et al. Circadian glucocorticoid oscillations are critical for learning-induced synaptic remodeling and maintenance. Nat Neurosci 2013 ; 16 : 698–705. [CrossRef] [PubMed] [Google Scholar]
- Yuste R. Dendritic spines. Cambridge MA : MIT Press, 2010. [CrossRef] [Google Scholar]
- Holsboer F, Ising M. Stress hormone regulation: biological role and translation into therapy. Annu Rev Psychol 2010 ; 61 : 81–109, C1–11. [CrossRef] [PubMed] [Google Scholar]
- Conway-Campbell BL, Pooley JR, Hager GL, Lightman SL. Molecular dynamics of ultradian glucocorticoid receptor action. Mol Cell Endocrinol 2012 ; 348 : 383–393. [CrossRef] [PubMed] [Google Scholar]
- Yehuda R. Post-traumatic stress disorder. N Engl J Med 2002 ; 346 : 108–114. [CrossRef] [PubMed] [Google Scholar]
- Green KN, Billings LM, Roozendaal B, et al. Glucocorticoids increase amyloid-beta and tau pathology in a mouse model of Alzheimer’s disease. J Neurosci 2006 ; 26 : 9047–9056. [CrossRef] [PubMed] [Google Scholar]
- Anacker C, Zunszain PA, Carvalho LA, Pariante CM. The glucocorticoid receptor: pivot of depression and of antidepressant treatment? Psychoneuroendocrinology 2011 ; 36 : 415–425. [CrossRef] [PubMed] [Google Scholar]
- Duman RS, Aghajanian GK. Synaptic dysfunction in depression: potential therapeutic targets. Science 2012 ; 338 : 68–72. [CrossRef] [PubMed] [Google Scholar]
- Prager EM, Johnson LR. Stress at the synapse: signal transduction mechanisms of adrenal steroids at neuronal membranes. Sci Signal 2009; 2 : re5. [CrossRef] [PubMed] [Google Scholar]
- McEwen BS. The ever-changing brain: cellular and molecular mechanisms for the effects of stressful experiences. Dev Neurobiol 2012 ; 72 : 878–890. [CrossRef] [PubMed] [Google Scholar]
- Malenka RC, Bear MF. LTP and LTD: an embarrassment of riches. Neuron 2004 ; 44 : 5–21. [CrossRef] [PubMed] [Google Scholar]
- Greer PL, Greenberg ME. From synapse to nucleus: calcium-dependent gene transcription in the control of synapse development and function. Neuron 2008 ; 59 : 846–860. [CrossRef] [PubMed] [Google Scholar]
- Kandel ER. The molecular biology of memory storage: a dialogue between genes and synapses. Science 2001 ; 294 : 1030–1038. [CrossRef] [PubMed] [Google Scholar]
- Lambert WM, Xu CF, Neubert TA, et al. BDNF-signaling rewrites the glucocorticoid transcriptome via glucocorticoid receptor phosphorylation. Mol Cell Biol 2013 ; 33 : 3700–3714. [CrossRef] [PubMed] [Google Scholar]
- Jeanneteau F, Chao MV. Are BDNF and glucocorticoid activities calibrated? Neuroscience 2013 ; 239 : 173–195. [CrossRef] [PubMed] [Google Scholar]
- Jeanneteau F, Deinhardt K, Miyoshi G, et al. The MAP kinase phosphatase MKP-1 regulates BDNF-induced axon branching. Nat Neurosci 2010 ; 13 : 1373–1379. [CrossRef] [PubMed] [Google Scholar]
- Duric V, Banasr M, Licznerski P, et al. A negative regulator of MAP kinase causes depressive behavior. Nat Med 2010 ; 16 : 1328–1332. [CrossRef] [PubMed] [Google Scholar]
- Duman RS. Neuronal damage and protection in the pathophysiology and treatment of psychiatric illness: stress and depression. Dialogues Clin Neurosci 2009 ; 11 : 239–255. [PubMed] [Google Scholar]
- Groc L, Chaouloff F. Axe corticotrope et plasticité de la communication neuronale : décryptage des mécanismes cellulaires. Med Sci (Paris) 2008 ; 24 : 776–778. [CrossRef] [EDP Sciences] [PubMed] [Google Scholar]
Liste des tableaux
Taux de renouvellement (mesuré sur trois jours) des épines dendritiques dans le cortex murin en réponse à différents changements de l’environnement. Le renouvellement des épines dendritiques augmente lorsque les souris sont maintenues dans un environnement enrichi (large cage + jouets) par rapport a un environnement basal (petite cage standard sans jouets). Les mêmes observations ont été constatées dans le cortex moteur lorsque les souris doivent acquérir des stratégies de coordination motrice pour ne pas chuter d’un Rotarod (3 séances d’apprentissage de 20 essais chacune). Souris adolescentes : âgées de 2 semaines à 1 mois maximum ; souris adultes : âgées de 4 mois au moins.
Liste des figures
|
Figure 1. Le pic circadien de glucocorticoïdes facilite l’apprentissage alors que le creux circadien est nécessaire à la consolidation de la mémoire. Cette phase critique de consolidation dure approximativement une semaine, pendant laquelle toute perturbation des oscillations de glucocorticoïdes a des conséquences néfastes sur la mémoire. En pointillé est illustré l’effet d’une injection de glucocorticoïdes exogènes sur les oscillations des taux de glucocorticoïdes endogènes. Pour limiter les effets du stress, les souris ont préalablement été habituées trois jours de suite sur un rotaRod non motorisé à la procédure qui sera utilisée lors de l’induction du pic de glucocorticoïdes. C’est l’accélération du rotaRod pendant 20 essais/jour à la même heure qui permet aux souris d’apprendre des stratégies de coordination et d’équilibre pour ne pas chuter. L’imagerie in vivo des neurones du cortex moteur lors du rappel de cette tâche après la phase de consolidation de la mémoire indique que la performance comportementale est proportionnelle à la survie des épines dendritiques formées au moment de l’apprentissage et à l’élimination d’épines formées avant l’apprentissage. Contrairement à l’apprentissage, qui est sensible au pic de glucocorticoïdes, la performance lors du rappel ne l’est pas. |
| Dans le texte | |
|
Figure 2. Conséquences d’une perturbation des rythmes circadiens de la sécrétion de glucocorticoïdes sur l’apprentissage et les bases structurales de la mémoire. L’apprentissage au moment du pic de glucocorticoïdes facilite l’acquisition et la consolidation de la mémoire et est corrélé à la formation d’un motif d’épines qui pourrait coder les bases structurales de l’engramme. L’absence chronique des pics de glucocorticoïdes empêche l’apprentissage et la consolidation de la mémoire qui est corrélé à un déficit des processus de formation et d’élimination des épines dendritiques. Au contraire, l’absence de creux circadiens due à un excès chronique de glucocorticoïdes produit des carences cognitives qui sont corrélées avec l’élimination accrue d’anciennes épines dendritiques et un taux de formation quasiment nul. |
| Dans le texte | |
|
Figure 3. Interférence des voies de signalisation des glucocorticoïdes et du BDNF via la phosphorylation des GR. Les glucocorticoïdes utilisent des mécanismes lents et rapides dépendants du GR pour augmenter le renouvellement d’épines dendritiques. La signalisation rapide des GR influence le remodelage du cytosquelette d’actine par la voie LIMK1-cofiline, alors que la signalisation lente des GR phosphorylés cible la transcription de certains gènes, dont celui qui code pour MKP1, une phosphatase influençant le remodelage du cytosquelette de microtubules. Ce mécanisme de signalisation est complexe et dépend du creux circadien du taux de glucocorticoïdes, et des récepteurs GR et MR. Il en résulterait un engramme permettant l’adaptation physiologique de ces réseaux neuronaux aux variations contextuelles. |
| Dans le texte | |
Les statistiques affichées correspondent au cumul d'une part des vues des résumés de l'article et d'autre part des vues et téléchargements de l'article plein-texte (PDF, Full-HTML, ePub... selon les formats disponibles) sur la platefome Vision4Press.
Les statistiques sont disponibles avec un délai de 48 à 96 heures et sont mises à jour quotidiennement en semaine.
Le chargement des statistiques peut être long.