")
")
| Issue |
Med Sci (Paris)
Volume 42, Number 5, Mai 2026
|
|
|---|---|---|
| Page(s) | 415 - 419 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/2026072 | |
| Published online | 29 May 2026 | |
PARP3 : un « couteau suisse » orchestrant la réparation de l’ADN, l’agressivité tumorale et la différenciation cellulaire
PARP3: a Swiss army knife orchestrating DNA repair, cancer aggressiveness and cell differentiation
Poly(ADP-ribosyl)ation et intégrité du génome, Laboratoire d’excellence Medalis, CNRS et université de Strasbourg UMR7242, Institut de recherche de l’école de biotechnologie de Strasbourg, Illkirch, France
*
This email address is being protected from spambots. You need JavaScript enabled to view it.
La poly(ADP-ribose) polymérase 3 (PARP3) est le troisième membre caractérisé de la famille des PARP, qui en contient 17. C’est une PARP ADN-dépendante, comme PARP1 et PARP2, dont elle se distingue cependant par son activité enzymatique, car elle catalyse la mono(ADP-ribosyl)ation, définie par l’addition d’un seul résidu ADP-ribose sur des protéines acceptrices ou sur l’ADN, tandis que PARP1 et PARP2 catalysent la poly(ADP-ribosyl)ation, c’est-à-dire l’addition de plusieurs unités d’ADPribose sur leurs cibles [1] (Figure 1). Dans l’espèce humaine, le gène PARP3 code deux isoformes issues d’un épissage alternatif du transcrit : une forme longue localisée au centrosome, et une forme courte nucléaire [2] (Figure 1). Chez la souris, seule la forme courte nucléaire est synthétisée.
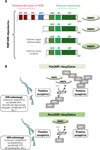 |
Figure 1 Structure modulaire des PARP ADNdépendantes et catalyse de l’ADP-ribosylation. A. Les PARP dépendantes de l’ADN (PARP1, PARP2 et PARP3) sont représentées avec leurs domaines : ZF (zinc finger), BRCT (BRCA1 C-terminus), WGR (Trp-Gly-Arg), HD (domaine hélicoïdal) et ART (ADP-ribosyltransférase). Il existe deux isoformes de PARP3 : une isoforme courte nucléaire (présente chez l’Homme et la souris) et une isoforme longue centrosomique (présente uniquement chez l’Homme) possédant 7 acides aminés supplémentaires à son extrémité N-terminale. B. Activées par les lésions de l’ADN (cassures, fourches de réplication bloquées, G4 quadruplex, extrémités d’ADN 5’P, etc.), PARP1 et PARP2 catalysent (avec NAD+ comme cosubstrat) la poly(ADPribosyl)ation pour greffer des chaînes d’ADPribose (ADPr) sur des protéines acceptrices, alors que PARP3 catalyse principalement la mono(ADP-ribosyl)ation. NAD+ : dinucléotide nicotinamide-adénine, forme oxydée ; N : nicotinamide. |
D’abord étudiée pour son rôle physiologique dans la réparation de l’ADN et la stabilité du génome, PARP3 a ensuite été associée à l’agressivité tumorale [3-5]. Les résultats de travaux de recherche récents permettent d’élargir le spectre fonctionnel de PARP3 au contrôle de la différenciation cellulaire dans un contexte physiologique [6, 7] (Figure 2).
 |
Figure 2 Rôles physiologiques de PARP3 et effet délétère dans un contexte tumoral : « le Yin et le Yang ». A. En contexte physiologique, PARP3 favorise la régénération tissulaire chez la planaire [16], contribue à la formation de la crête neurale au cours du développement du poissonzèbre [15], participe à la neurogenèse [7], et joue un rôle essentiel dans la myogenèse et le fonctionnement du muscle squelettique chez la souris [6]. B. Dans un contexte tumoral, PARP3 est un acteur clé de l’agressivité de la tumeur [3-5]. C. Les rôles de PARP3 dans la réparation des lésions de l’ADN [8-10, 13], la commutation isotypique des immunoglobulines (Ig) [12], la ségrégation des télomères [9], la division mitotique [9], et les réarrangements chromosomiques [13], concernent aussi bien la physiologie que la pathologie. |
PARP3, acteur polyvalent de la réparation de l’ADN
PARP3 est impliquée dans la réparation des cassures simple-brin et doublebrin de l’ADN. Concernant les cassures simple-brin, PARP3 agit préférentiellement dans le nucléosome1, où elle catalyse la mono-ADP-ribosylation des histones (par exemple H2B) : elle joue donc un rôle de senseur de cassures simplebrin dans un contexte chromatinien. Dans le modèle aviaire DT402, l’absence de PARP3 ralentit la cinétique de réparation des cassures simple-brin induites par l’irradiation g [8]. PARP3 joue un rôle déterminant dans le choix du mode de réparation des cassures double-brin, qu’elle oriente vers la jonction d’extrémités non homologues « classique » (canonical DNA non-homologous endjoining, c-NHEJ). En catalysant l’ADPribosylation du complexe protéique dimérique Ku70/Ku80, acteur central de la voie c-NHEJ, elle favorise le recrutement de ce complexe ou son maintien sur les cassures, elle active l’étape de ligation des extrémités d’ADN en coopération avec le facteur APLF (aprataxin-and-PNK-like), et elle limite la résection des extrémités d’ADN, une étape clé de la recombinaison homologue ou du mode « alternatif » de réparation des cassures par jonction des extrémités (alternative end-joining, alt-EJ) [9-11]. PARP3 est impliquée dans la réparation des cassures double-brin physiologiques de l’ADN provoquées par la cytidine désaminase « induite par activation » (activation-induced cytidine deaminase, AID) lors de la commutation de classe des immunoglobulines, dans le maintien de l’intégrité des télomères, et dans la suppression de structures d’ADN « non canoniques » telles que les quadruplex de guanine (G4) lorsqu’ils sont induits chimiquement par des ligands de G4 [9, 12, 13].
PARP3, acteur central de l’agressivité tumorale
Les résultats de plusieurs travaux de recherche ont montré la contribution majeure de PARP3 dans l’agressivité tumorale, dans des contextes où PARP1 semble moins abondante ou moins active. Dans le cancer du sein, PARP3 est plus abondante dans des lignées de cellules tumorales dont le phénotype est mésenchymateux et invasif que dans des lignées épithéliales moins mobiles. Sa présence reste peu détectable dans une lignée de cellules mammaires normales [4]. En accord avec ces constats, PARP3 contribue aux caractéristiques des tumeurs agressives : elle facilite la transition épithélio-mésenchymateuse induite par TGFb et les dérivés radicalaires de l’oxygène, promeut le renouvellement des cellules souches cancéreuses, et active la voie de signalisation oncogénique RICTOR (rapamycin-insensitive companion of mTOR) / mTORC2. La déplétion, l’inactivation, ou l’inhibition pharmacologique de PARP3 réduit l’agressivité tumorale [3, 4]. Dans le modèle du glioblastome, l’interaction de PARP3 avec la méthyltransférase G9a limite son activité de synthèse de la marque répressive de la chromatine sur l’histone H3 (H3K9me2) pour permettre l’expression de gènes impliqués dans l’adhérence cellulaire ou dans la réponse à l’hypoxie. L’absence de PARP3 sensibilise les cellules tumorales à l’hypoxie et augmente leur sensibilité aux médicaments anticancéreux de la famille des vinca-alcaloïdes3 [5]. PARP3 est donc une cible prometteuse d’oncothérapie, en particulier contre les cancers agressifs. La production de molécules synthétiques de la famille des quinazolinones, suivie d’un profilage enzymatique sur les PARP, a permis d’identifier un inhibiteur compétitif du site de liaison de PARP3 à son cosubstrat NAD+ (dinucléotide nicotinamide-adénine, forme oxydée) : la molécule ME0328 [14]. Bien que PARP3 soit inhibée par ME0328 environ sept fois plus que ne l’est PARP1 in vitro, cette molécule n’est active dans les cellules qu’à des concentrations submicromolaires. Il est donc nécessaire d’identifier ou de synthétiser des molécules présentant une activité inhibitrice de PARP3 plus sélective ainsi qu’une meilleure stabilité métabolique, avant d’envisager une utilisation thérapeutique.
PARP3, acteur de la différenciation cellulaire physiologique
La découverte d’une implication de PARP3 dans l’induction de la transition épithélio-mésenchymateuse de cellules cancéreuses [4], considérée comme un processus de différenciation cellulaire transitoire et réversible, et de son rôle dans la spécification de la crête neurale embryonnaire chez le poisson-zèbre [15], ont motivé la recherche d’un rôle physiologique dans la différenciation cellulaire.
PARP3 a ainsi été impliquée dans la différenciation de cellules souches et progénitrices neurales du cerveau en astrocytes, dans un modèle murin. D’un point de vue mécanistique, PARP3 modère la production de dérivés radicalaires de l’oxygène catalysée par la NADPH oxydase NOX4, en limitant la translocation vers le noyau du facteur de transcription NF-κB impliqué dans la synthèse de cette oxydase. L’absence de PARP3 déclenche localement un stress oxydant caractérisé par une surproduction de dérivés radicalaires de l’oxygène, responsable de l’oxydation et de la dégradation du complexe RICTOR/mTORC2 par l’ubiquitine ligase FBXW7 (Skp cullin F-box/WD repeat-containing protein 7), spécifiquement dans le striatum4. Sur le plan fonctionnel, cela se traduit par la persistance d’astrocytes immatures et une vulnérabilité accrue à l’hypoxie cérébrale [7].
Nous avons par ailleurs montré que la présence de PARP3 est nécessaire à la différenciation de cellules souches musculaires (cellules satellites) en myotubes et au maintien de l’intégrité du muscle squelettique dans un modèle murin. Dans la lignée de cellules myoblastiques murines C2C12, un modèle de référence pour l’étude de la différenciation musculaire in vitro, l’inactivation de Parp3 perturbe sévèrement la différenciation myogénique, qui s’arrête au stade de myocytes immatures caractérisés par une diminution de l’expression de marqueurs myogéniques (MyoD, MyoG), un cytosquelette d’actine désorganisé, et l’apparition de mitochondries dégénérées et de vacuoles d’autophagie. Bien que les conséquences d’une suppression de PARP3 chez la souris apparaissent plus modestes, les altérations fonctionnelles demeurent. Elles se traduisent par une faiblesse du muscle squelettique Tibialis anterior chez les jeunes adultes, l’accumulation de myotubes à noyaux centraux associée à une augmentation de l’expression de gènes impliqués dans la régénération musculaire, et une reprogrammation de la composition de ce muscle en fibres contractiles avec une diminution du nombre des fibres à contraction rapide (fibres IIa) au profit de fibres à contraction lente (fibres IIx). Les cellules souches satellites de ces souris conservent une capacité de différenciation myogénique, mais affectée par une diminution de l’expression de gènes impliqués dans la différenciation, tels que MyoD. D’un point de vue mécanistique, nos résultats indiquent l’existence d’une interaction entre PARP3 et l’histone méthyltransférase EZH2, sous-unité catalytique du complexe PRC2 (polycomb repressive complex 2) qui catalyse la marque épigénétique répressive H3K27me3 sur l’histone H3. PARP3 catalyse l’ADP-ribosylation de EZH2 et exerce un rôle modulateur binaire sur l’enrichissement de la marque H3K27me3 au contact de gènes clés de la différenciation : alors que PARP3 favorise l’abondance de cette marque répressive de la transcription pour les gènes Igfbp3 (Insulin-like growth factor-binding protein 3) et Epha7 (Ephrin type-A receptor 7), elle en réprime au contraire l’accumulation pour le gène MyoD [6].
Ces deux découvertes soulignent l’intérêt d’identifier des molécules thérapeutiques activatrices de PARP3, qui seraient utilisables contre l’ischémie cérébrale, des maladies neurodégénératives, ou des maladies musculaires.
Perspectives
Les résultats des recherches actuelles mettent en évidence les fonctions multiples de la protéine PARP3 et son rôle ambivalent en partie dépendant du contexte : elle est promotrice de l’agressivité tumorale, tout en jouant un rôle essentiel dans la réparation des cassures de l’ADN et en contribuant à la différenciation cellulaire physiologique dans la myogenèse et la neurogenèse (Figure 2). Toutefois, une cohérence fonctionnelle semble émerger de ses propriétés moléculaires, de son rôle dans la maintenance de l’intégrité du génome, et de son interaction avec des modulateurs ou des composants de la chromatine (G9a, EZH2, H2B) afin de contrôler la plasticité cellulaire (comme en témoigne par exemple son implication dans la transition épithélio-mésenchymateuse de cellules cancéreuses). L’ensemble des résultats révèle une logique structurante : la capacité de PARP3 à intégrer les signaux de l’ADN et de la chromatine pour orienter le destin cellulaire en fonction du contexte physiologique ou pathologique. Une analyse approfondie de ses mécanismes d’action ainsi que de ses partenaires et cibles moléculaires reste à faire. À cela s’ajoute la nécessité d’études translationnelles visant à identifier des molécules inhibitrices ou activatrices présentant un potentiel thérapeutique, notamment contre le cancer, les maladies neurodégénératives, ou les maladies musculaires.
Liens d’intérêt
Les auteurs déclarent ne pas avoir de lien d’intérêt.
Références
- Dasovich M, Leung AKL. PARPs and ADP-ribosylation: Deciphering the complexity with molecular tools. Mol Cell 2023 ; 83 : 1552-72. [Google Scholar]
- Augustin A, Spenlehauer C, Dumond H, et al. PARP-3 localizes preferentially to the daughter centriole and interferes with the G1/S cell cycle progression. J Cell Sci 2003 ; 116 : 1551-62. [Google Scholar]
- Beck C, Rodriguez-Vargas JM, Boehler C, et al. PARP3, a new therapeutic target to alter Rictor/mTORC2 signaling and tumor progression in BRCA1-associated cancers. Cell Death Differ 2019 ; 26 : 1615-30. [Google Scholar]
- Karicheva O, Rodriguez-Vargas JM, Wadier N, et al. PARP3 controls TGFβ and ROS driven epithelial-to-mesenchymal transition and stemness by stimulating a TG2-Snail-E-cadherin axis. Oncotarget 2016 ; 7 : 64109-23. [Google Scholar]
- Nguekeu-Zebaze L, Hanini N, Noll A, et al. PARP3 supervises G9a-mediated repression of adhesion and hypoxia-responsive genes in glioblastoma cells. Sci Rep 2022 ; 12 : 15534. [Google Scholar]
- Yildirim Z, Noll A, Martin-Hernandez K, et al. Parp3 assists muscle function and skeletal muscle differentiation by selectively adjusting H3K27me3 enrichment. iScience 2025 ; 28 : 112267. [Google Scholar]
- Rodriguez-Vargas JM, Martin-Hernandez K, Wang W, et al. Parp3 promotes astrocytic differentiation through a tight regulation of Nox4-induced ROS and mTorc2 activation. Cell Death Dis 2020 ; 11 : 954. [Google Scholar]
- Grundy GJ, Polo LM, Zeng Z, et al. PARP3 is a sensor of nicked nucleosomes and monoribosylates histone H2B(Glu2). Nat Commun 2016 ; 7 : 12404. [Google Scholar]
- Boehler C, Gauthier LR, Mortusewicz O, et al. Poly(ADP-ribose) polymerase 3 (PARP3), a newcomer in cellular response to DNA damage and mitotic progression. Proc Natl Acad Sci USA 2011 ; 108 : 2783-8. [Google Scholar]
- Beck C, Boehler C, Barbat JG, et al. PARP3 affects the relative contribution of homologous recombination and nonhomologous end-joining pathways. Nucleic Acids Res 2014 ; 42 : 5616-32. [Google Scholar]
- Rulten SL, Fisher AEO, Robert I, et al. PARP-3 and APLF function together to accelerate nonhomologous end-joining. Mol Cell 2011 ; 41 : 33-45. [Google Scholar]
- Robert I, Gaudot L, Rogier M, et al. Parp3 negatively regulates immunoglobulin class switch recombination. PLoS Genet 2015 ; 11 : e1005240. [Google Scholar]
- Day TA, Layer JV, Cleary JP, et al. PARP3 is a promoter of chromosomal rearrangements and limits G4 DNA. Nat Commun 2017 ; 8 : 15110. [Google Scholar]
- Lindgren AE, Karlberg T, Thorsell AG, et al. A PARP inhibitor with selectivity toward ADP-ribosyltransferase ARTD3/PARP3. ACS Chem Biol 2013 ; 8 : 1698-703. [Google Scholar]
- Rouleau M, Saxena V, Rodrigue A, et al. A key role for poly(ADP-ribose) polymerase 3 in ectodermal specification and neural crest development. PLoS One 2011 ; 6 : e15834. [Google Scholar]
- Barghouth PG, Karabinis P, Venegas A, et al. Poly(ADP-ribose) polymerase-3 regulates regeneration in planarians. Int J Mol Sci 2020 ; 21 : 875. [Google Scholar]
Complexe comportant un segment d’ADN de 146 ou 147 paires de nucléotides, enroulé autour d’un cœur formé de protéines (histones). Chez les eucaryotes, le nucléosome constitue l’unité d’organisation de la chromatine (source : Wikipédia).
Lignée de lymphoblastes B de poulet provenant d’un lymphome de la bourse de Fabricius (organe lymphoïde chez les oiseaux).
Alcaloïdes isolés de différentes pervenches tropicales, surtout Catharanthus roseus (appelé auparavant Vinca rosea, d’où le nom de ces alcaloïdes). Ils représentent la plus ancienne famille d’agents anticancéreux ciblant les microtubules (source : Wikipédia).
Noyau cérébral sous cortical majeur de la boucle mésocorticolimbique, impliqué dans les comportements moteurs, cognitifs et émotionnels.
© 2026 médecine/sciences – Inserm
 Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l’utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l’utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Liste des figures
|
Figure 1 Structure modulaire des PARP ADNdépendantes et catalyse de l’ADP-ribosylation. A. Les PARP dépendantes de l’ADN (PARP1, PARP2 et PARP3) sont représentées avec leurs domaines : ZF (zinc finger), BRCT (BRCA1 C-terminus), WGR (Trp-Gly-Arg), HD (domaine hélicoïdal) et ART (ADP-ribosyltransférase). Il existe deux isoformes de PARP3 : une isoforme courte nucléaire (présente chez l’Homme et la souris) et une isoforme longue centrosomique (présente uniquement chez l’Homme) possédant 7 acides aminés supplémentaires à son extrémité N-terminale. B. Activées par les lésions de l’ADN (cassures, fourches de réplication bloquées, G4 quadruplex, extrémités d’ADN 5’P, etc.), PARP1 et PARP2 catalysent (avec NAD+ comme cosubstrat) la poly(ADPribosyl)ation pour greffer des chaînes d’ADPribose (ADPr) sur des protéines acceptrices, alors que PARP3 catalyse principalement la mono(ADP-ribosyl)ation. NAD+ : dinucléotide nicotinamide-adénine, forme oxydée ; N : nicotinamide. |
| Dans le texte | |
|
Figure 2 Rôles physiologiques de PARP3 et effet délétère dans un contexte tumoral : « le Yin et le Yang ». A. En contexte physiologique, PARP3 favorise la régénération tissulaire chez la planaire [16], contribue à la formation de la crête neurale au cours du développement du poissonzèbre [15], participe à la neurogenèse [7], et joue un rôle essentiel dans la myogenèse et le fonctionnement du muscle squelettique chez la souris [6]. B. Dans un contexte tumoral, PARP3 est un acteur clé de l’agressivité de la tumeur [3-5]. C. Les rôles de PARP3 dans la réparation des lésions de l’ADN [8-10, 13], la commutation isotypique des immunoglobulines (Ig) [12], la ségrégation des télomères [9], la division mitotique [9], et les réarrangements chromosomiques [13], concernent aussi bien la physiologie que la pathologie. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.