")
")
| Issue |
Med Sci (Paris)
Volume 37, Number 4, Avril 2021
|
|
|---|---|---|
| Page(s) | 324 - 326 | |
| Section | Le Magazine | |
| DOI | https://doi.org/10.1051/medsci/2021026 | |
| Published online | 28 April 2021 | |
Réparation tissulaire : le rôle clé de l’annexine A1 dans le contrôle de la réaction inflammatoire
Tissue repair: Key role of annexin A1 in the control of inflammatory response
Institut NeuroMyoGène, Université Claude Bernard Lyon 1, CNRS UMR5310, Inserm U1217, Université Lyon, 8 avenue Rockfeller, 69008 Lyon, France.
* This email address is being protected from spambots. You need JavaScript enabled to view it.
** This email address is being protected from spambots. You need JavaScript enabled to view it.
Une réponse inflammatoire efficace est une composante nécessaire de la réaction aux infections et aux lésions tissulaires. L’homéostasie tissulaire exige que ce processus se déroule de façon appropriée et cesse dans une certaine temporalité [1]. Une réponse inflammatoire d’intensité excessive, ou qui se prolonge anormalement et devient chronique, caractérise diverses maladies, telles que le sepsis, le diabète ou la polyarthrite rhumatoïde [1].
Le muscle strié squelettique : un modèle d’étude de la réparation tissulaire
Le muscle strié squelettique est un tissu contractile principalement composé de cellules multinucléées : les fibres musculaires. Il constitue un excellent modèle d’étude de la réparation tissulaire car il est capable de se régénérer complètement grâce à des cellules souches localisées à la périphérie des fibres musculaires : les cellules satellites [2]. A l’état basal, ces cellules sont quiescentes. Après une lésion, elles s’activent, prolifèrent, se différencient, puis fusionnent pour former de nouvelles fibres. Ce processus de régénération dépend fortement de l’interaction des cellules satellites avec d’autres types de cellules, parmi lesquelles les cellules de l’inflammation (i.e., granulocytes neutrophiles et monocytes), qui infiltrent rapidement le tissu endommagé [3]. Les monocytes se différencient en macrophages pro-inflammatoires, qui phagocytent les fibres musculaires endommagées et favorisent la prolifération des cellules satellites [4]. Ils changent ensuite de phénotype pour devenir des macrophages anti-inflammatoires stimulant la fusion des cellules myogéniques, et favorisent ainsi la formation des nouvelles fibres musculaires et la régénération du tissu musculaire [5]. Ce changement de statut inflammatoire, en partie contrôlé par la protéine enzymatique AMPK (AMP-activated kinase), est un évènement crucial du processus de régénération [6], et son altération est associée à des maladies telles que les myopathies dégénératives [7].
L’AMPK n’est pas uniquement un senseur énergétique
À l’instar des HIF (hypoxia-inducible factors) et des PPAR (peroxisome proliferator-activated receptors), l’AMPK est un senseur énergétique de la cellule [8]. Cette protéine hétérotrimérique est composée d’une sous-unité catalytique (a) et de deux sous-unités régulatrices (b et g). Elle est activée par les kinases LKB1 (liver kinase B1) et CaMKKb (calcium/calmodulin-dependent protein kinase kinase b), ainsi que par la variation des concentrations cellulaires d’AMP, d’ATP, d’ADP ou de glycogène. Son rôle dans différents processus métaboliques est bien documenté. L’AMPK est aussi impliquée dans le cycle et la polarité cellulaires [8]. Récemment, son implication dans la modulation des processus inflammatoires a été mise en évidence dans différents contextes : infection, inflammation aiguë non infectieuse, et inflammation chronique. Nous avons précédemment montré que l’AMPK agit comme un signal intracellulaire permettant l’acquisition, par les macrophages, d’un phénotype anti-inflammatoire favorisant la résolution de l’inflammation et la régénération du muscle squelettique après une lésion aiguë ou chronique [6]. En revanche, l’élément extracellulaire déclencheur de cette transformation phénotypique des macrophages restait jusqu’à présent mal compris.
L’acquisition du statut inflammatoire des macrophages est contrôlée par la voie de signalisation annexine1/FPR2/AMPK
L’annexine A1 est une protéine produite principalement par les leucocytes et les cellules stromales. Elle est sécrétée dans le milieu extracellulaire et se lie au récepteur FPR2 (formyl peptide receptor 2), un récepteur membranaire couplé aux protéines G [9]. L’annexine A1 est un acteur majeur de la résolution de l’inflammation par son implication dans le contrôle de l’apoptose des granulocytes neutrophiles, le recrutement des monocytes et la phagocytose des macrophages [10]. En outre, elle promeut l’acquisition par les macrophages d’un phénotype anti-inflammatoire dans des modèles in vitro d’arthrite rhumatoïde [11] et de croissance tumorale [12].
Afin de caractériser le mécanisme par lequel l’annexine A1 intervient dans ce changement phénotypique des macrophages, des modèles de différenciation in vitro de macrophages dérivés de la moelle osseuse murine ou de sang humain ont été utilisés [13]. Ces expériences ont permis de montrer que l’annexine A1 favorise l’acquisition du phénotype anti-inflammatoire par les macrophages en se liant au récepteur FPR2 et en induisant la phosphorylation de la protéine AMPK. En effet, des macrophages dans lesquels le gène codant le récepteur FPR2 ou celui codant la sous-unité α1 de l’AMPK a été inactivé, ne sont plus sensibles à l’action de l’annexine A1. Ces résultats ont été confirmés in vivo dans un modèle murin de lésion du muscle tibialis anterior induite par une injection de cardiotoxine (une toxine de venin de serpent).En effet, une proportion significative des macrophages pro-inflammatoires ne produisant plus d’annexine A1 ou n’exprimant plus son récepteur FPR2 ont perdu la capacité d’acquérir les propriétés anti-inflammatoires indispensables à la résolution de l’inflammation. La perte de ce changement phénotypique des macrophages est associée à un défaut de régénération musculaire.
L’annexine A1 : un acteur clé de la régénération du muscle strié squelettique
En utilisant le même modèle murin, nous avons montré qu’au cours de la régénération musculaire, l’annexine A1 est produite dès le début de la réponse inflammatoire, d’abord par les granulocytes neutrophiles, puis par les macrophages pro-inflammatoires [13]. Remarquablement, l’inactivation du gène codant l’annexine A1 (AnxA1), ou celle du gène Fpr2 codant son récepteur, chez la souris, altère la régénération musculaire après une blessure. L’inactivation du gène AnxA1 restreinte aux cellules myéloïdes a le même effet, confirmant leur rôle dans la production de l’annexine A1 nécessaire à la régénération musculaire. De plus, des expériences in vitro ont permis de confirmer que les effets de l’annexine A1 au cours de la régénération musculaire sont principalement dus aux macrophages. En effet, alors qu’un traitement direct des cellules myogéniques avec l’annexine A1 est sans effet sur leur fusion, celle-ci est stimulée par le milieu conditionné issu de macrophages traités avec l’annexine A1. Cet effet est perdu lorsque le milieu conditionné est issu de macrophages dans lesquels le gène codant la sous-unité α1 de l’AMPK a été inactivé, confirmant le rôle prépondérant de l’AMPK dans ce processus.
Les résultats de cette étude effectuée sur un modèle de lésion musculaire ont montré l’importance de l’activation de la voie de signalisation ANXA1/FPR2/AMPK dans les macrophages, via une production paracrine et autocrine de l’annexine A1, pour la résolution de l’inflammation, permettant la réparation tissulaire après une lésion (Figure 1). Ils ouvrent ainsi la voie à une recherche de nouveaux traitements contre des maladies caractérisées par une inflammation excessive ou chronique, telles que le sepsis ou les myopathies dégénératives.
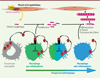 |
Figure 1. Modèle pour l’action de l’annexine A1 au cours de la régénération du muscle strié squelettique. Après une lésion musculaire, les cellules souches musculaires (cellules satellites) sont activées. Parallèlement, le tissu endommagé est infiltré par les granulocytes neutrophiles et les monocytes. Ces derniers se différencient en macrophages pro-inflammatoires favorisant la prolifération des cellules satellites. L’annexine A1 (ANX A1), produite à la fois par les granulocytes neutrophiles et les macrophages pro-inflammatoires, se fixe sur le récepteur FPR2 présent sur les macrophages pro-inflammatoires et induit la phosphorylation de la protéine kinase AMPK (AMP-activated kinase). L’enzyme AMPK, ainsi activée, promeut l’acquisition du phénotype anti-inflammatoire par les macrophages, une étape cruciale pour stimuler la fusion des cellules myogéniques et la formation des nouvelles fibres musculaires. |
Liens d’intérêt
Les auteurs déclarent n’avoir aucun lien d’intérêt concernant les données publiées dans cet article.
Références
- Perretti M, Leroy X, Bland EJ, et al. Resolution pharmacology: opportunities for therapeutic innovation in inflammation. Trends Pharmacol Sci 2015 ; 36 : 737–755. [CrossRef] [PubMed] [Google Scholar]
- Yin H, Price F, Rudnicki MA. Satellite cells and the muscle stem cell niche. Physiol Rev 2013 ; 93 : 23–67. [Google Scholar]
- Chazaud B.. Macrophages: supportive cells for tissue repair and regeneration. Immunobiology 2014 ; 219 : 172–178. [CrossRef] [PubMed] [Google Scholar]
- Saclier M, Cuvellier S, Magnan M, et al. Monocyte/macrophage interactions with myogenic precursor cells during skeletal muscle regeneration. FEBS J 2013 ; 280 : 4118–4130. [CrossRef] [PubMed] [Google Scholar]
- Arnold L, Henry A, Poron F, et al. Inflammatory monocytes recruited after skeletal muscle injury switch into antiinflammatory macrophages to support myogenesis. J Exp Med 2007 ; 204 : 1057–1069. [CrossRef] [PubMed] [Google Scholar]
- Mounier R, Theret M, Arnold L, et al. AMPKa1 regulates macrophage skewing at the time of resolution of inflammation during skeletal muscle regeneration. Cell Metab 2013 ; 18 : 251–264. [CrossRef] [PubMed] [Google Scholar]
- Juban G, Saclier M, Yacoub-Youssef H, et al. AMPK Activation regulates LTBP4-dependent TGF-β1 secretion by pro-inflammatory macrophages and controls fibrosis in Duchenne muscular dystrophy. Cell Rep 2018 ; 25 : 2163–2176. [CrossRef] [PubMed] [Google Scholar]
- Hardie DG. AMP-activated protein kinase: an energy sensor that regulates all aspects of cell function. Genes Dev 2011 ; 25 : 1895–1908. [CrossRef] [PubMed] [Google Scholar]
- Perretti M, Dalli J. Exploiting the annexin A1 pathway for the development of novel anti-inflammatory therapeutics. Br J Pharmacol 2009 ; 158 : 936–946. [CrossRef] [PubMed] [Google Scholar]
- Lim LH, Pervaiz S. Annexin 1: the new face of an old molecule. FASEB J 2007 ; 21 : 968–975. [CrossRef] [PubMed] [Google Scholar]
- Rhys HI, Dell’Accio F, Pitzalis C, et al. Neutrophil microvesicles from healthy control and rheumatoid arthritis patients prevent the inflammatory activation of macrophages. EBioMedicine 2018 ; 29 : 60–69. [CrossRef] [PubMed] [Google Scholar]
- Moraes LA, Kar S, Foo SJ, et al. Annexin-A1 enhances breast cancer growth and migration by promoting alternative macrophage polarization in the tumour microenvironment. Sci Rep 2017 ; 7 : 17925. [CrossRef] [PubMed] [Google Scholar]
- McArthur S, Juban G, Gobbetti T, et al. Annexin A1 drives macrophage skewing to accelerate muscle regeneration through AMPK activation. J Clin Invest 2020; 130 : 1156–67. [CrossRef] [PubMed] [Google Scholar]
© 2021 médecine/sciences – Inserm
 Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l'utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l'utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Liste des figures
|
Figure 1. Modèle pour l’action de l’annexine A1 au cours de la régénération du muscle strié squelettique. Après une lésion musculaire, les cellules souches musculaires (cellules satellites) sont activées. Parallèlement, le tissu endommagé est infiltré par les granulocytes neutrophiles et les monocytes. Ces derniers se différencient en macrophages pro-inflammatoires favorisant la prolifération des cellules satellites. L’annexine A1 (ANX A1), produite à la fois par les granulocytes neutrophiles et les macrophages pro-inflammatoires, se fixe sur le récepteur FPR2 présent sur les macrophages pro-inflammatoires et induit la phosphorylation de la protéine kinase AMPK (AMP-activated kinase). L’enzyme AMPK, ainsi activée, promeut l’acquisition du phénotype anti-inflammatoire par les macrophages, une étape cruciale pour stimuler la fusion des cellules myogéniques et la formation des nouvelles fibres musculaires. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.