")
")
| Issue |
Med Sci (Paris)
Volume 35, Number 12, Décembre 2019
Anticorps monoclonaux en thérapeutique
|
|
|---|---|---|
| Page(s) | 1106 - 1112 | |
| Section | Les nouveaux formats d’anticorps | |
| DOI | https://doi.org/10.1051/medsci/2019223 | |
| Published online | 06 January 2020 | |
Améliorer le ciblage tissulaire des anticorps thérapeutiques par de nouveaux formats
L’exemple de la barrière hémato-encéphalique
New formats for improving brain drug delivery of antibodies: the blood-brain barrier case
1
Institut Pasteur, Plateforme d’ingénierie des anticorps, Paris, France
2
Sanofi, Département des Maladies Rares et Neurologiques, Groupe Barrières Cérébrales, Paris, France
3
Université Paris Descartes, Faculté de Pharmacie de Paris et Inserm UMRS-1144, Barrière hémato-encéphalique: Physiopathologie et Thérapie, Paris, France
* This email address is being protected from spambots. You need JavaScript enabled to view it.
Résumé
De nombreuses pathologies cérébrales neurodégénératives ou tumorales devraient pourvoir bénéficier des progrès thérapeutiques impressionnants des anticorps médicaments. Malheureusement, en raison de leur très faible passage dans le cerveau, de nombreux développements cliniques d’anticorps dont la cible thérapeutique se situe dans le parenchyme cérébral ont été arrêtés par manque d’efficacité. La barrière hémato-encéphalique (BHE), douée de propriétés extrêmement sélectives et restrictives, est à l’origine de la faible pénétration cérébrale des molécules de haute masse moléculaire, telles que les anticorps thérapeutiques. L’objectif de cette revue est de présenter les propriétés de la BHE et les dernières avancées dans le domaine de l’ingénierie de nouveaux formats d’anticorps susceptibles d’améliorer leur passage intracérébral.
Abstract
Many neurodegenerative or tumor brain pathologies should be able to benefit from the impressive medicinal advances that represent therapeutic antibodies. Unfortunately, many failures have been observed with antibodies whose targets are in the brain parenchyma due to their very low brain distribution. The blood-brain barrier (BBB) that exhibits extremely selective and restrictive properties is responsible for the low brain penetration of high-molecular mass molecules including therapeutic antibodies. The objective of this article is to present the properties of the BBB and the latest advances in the engineering of new antibody formats to possibly improve their brain distribution.
© 2019 médecine/sciences – Inserm
 Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l'utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l'utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Bien que de très nombreux anticorps thérapeutiques aient été mis sur le marché depuis une décennie pour traiter principalement des cancers et des maladies inflammatoires, seuls 5 % d’entre eux sont indiqués dans des maladies du système nerveux central (SNC). En France, le natalizumab et l’alemtuzumab sont indiqués dans la sclérose en plaque (SEP) et le bévacizumab vient d’obtenir l’autorisation de mise sur le marché (AMM) des autorités américaines de la FDA (food and drug administration) dans le traitement d’une tumeur cérébrale, le glioblastome. Si l’on s’intéresse au mécanisme d’action de ces anticorps thérapeutiques dans ces maladies du SNC, on s’aperçoit qu’il ne repose pas sur leur passage du sang vers le cerveau mais en bloquant dans le sang soit l’extravasation cérébrale des leucocytes dans la SEP, soit en inhibant la fixation du VEGF (vascular endothelial growth factor) à son récepteur, ce facteur de croissance agissant positivement sur la croissance tumorale dans le glioblastome. De nombreux anticorps à visée thérapeutique ont été, ou sont encore actuellement testés dans des essais cliniques dans diverses pathologies et notamment celles du SNC comme la maladie d’Alzheimer (MA). Malheureusement, la plupart de ces anticorps thérapeutiques ont vu leur développement arrêté après les essais de phases II ou III par manque d’efficacité ou du fait de la survenue d’effets secondaires au niveau cérébral, comme des vasculopathies ayant entraîné des microhémorragies cérébrales et des œdèmes vasogéniques. La raison principale de l’absence sur le marché d’anticorps thérapeutiques ayant pour cibles des protéines du parenchyme cérébral est le maintien dans ces pathologies de barrières cérébrales intactes, limitant ainsi très efficacement leur entrée dans le cerveau. On estime ainsi la fraction biodisponible pour le tissu cérébral d’anticorps administrés par voie intraveineuse à moins de 0,1 % de la dose injectée [1], ce qui est la plupart du temps insuffisant pour espérer un effet thérapeutique. Le parenchyme cérébral est en effet très efficacement protégé des molécules et cellules sanguines circulantes par deux barrières entre le sang et le tissu cérébral: (1) la barrière hémato-liquidienne (BHL), entre le sang et le liquide céphalo-rachidien (LCR), au niveau des espaces méningés sous-arachnoïdiens et des plexus choroïdes localisés au niveau des ventricules cérébraux, et (2) la barrière hémato-encéphalique (BHE) qui sépare le parenchyme cérébral du sang au niveau de la plupart des structures cérébrales de la substance blanche et grise. Bien que la BHL soit environ 100 fois plus perméable que la BHE au passage passif de composés quelle que soit leur taille moléculaire, sa surface d’échange est 100 fois plus faible [2]. Les molécules qui entrent plus facilement dans le LCR à travers la BHL que dans le parenchyme cérébral à travers la BHE ont, de plus, une capacité de diffusion très limitée du LCR vers le parenchyme cérébral. On estime la distance de diffusion d’une molécule biologique depuis le LCR vers le parenchyme cérébral à moins de 1 mm avec une concentration qui diminue de façon logarithmique à partir de la surface du LCR. Même si l’on s’affranchit du passage d’un médicament du sang dans le LCR en l’administrant directement dans le LCR par voie intrathécale, le fort taux de renouvellement (3 à 4 fois par jour) des 140 ml de LCR chez l’homme et sa convection (mouvement des fluides cérébraux) des ventricules vers les sinus veineux des espaces sous-arachnoïdiens, entraînent une sortie rapide du médicament du LCR vers le sang. Ainsi, même si l’administration d’un médicament biologique par voie intraveineuse permet un meilleur passage de celui-ci dans le LCR à travers la BHL, la faible surface d’échange de cette barrière et les mécanismes rapides de clairance du LCR ne permettent pas une diffusion intracérébrale satisfaisante. Afin d’espérer une efficacité thérapeutique des anticorps ayant pour cibles des protéines intracérébrales dans des pathologies du SNC à BHE conservée, leur passage à travers la BHE doit donc être considérablement augmenté. Nous présentons dans cette revue les progrès actuels à la fois dans la compréhension des mécanismes moléculaires de transport à travers la BHE et dans les possibilités d’ingénierie de nouveaux formats d’anticorps ciblant la BHE qui pourraient permettre de pallier ces problèmes de distribution cérébrale des médicaments biologiques.
Propriétés de la BHE
La barrière hémato-encéphalique (BHE) est une structure essentielle du cerveau puisqu’elle maintient l’homéostasie indispensable au bon déroulement des différentes activités cérébrales. Cette barrière est constituée d’une couche continue de cellules endothéliales jointives reposant sur une lame basale incrustée de péricytes1 et solidement gainée par les pieds des astrocytes qui recouvrent pratiquement totalement la surface des microvaisseaux cérébraux (Figure 1) [3].
 |
Figure 1. Structure histologique de la barrière hémato-encéphalique (BHE). A. Schéma d’un capillaire cérébral en coupe transversale montrant les différentes structures cellulaires composant la BHE. B. Visualisation par microscopie confocale d’un vaisseau cérébral dans le cortex chez le rat. Les astrocytes sont révélés par un immunomarquage de la GFAP (glial fibrillary acidic protein) (en jaune) et la membrane luminale des cellules endothéliales par un immunomarquage de la P-glycoprotéine (en magenta). Les noyaux cellulaires (cellules endothéliales, cellules murales, neurones, cellules gliales) sont marqués par un intercalant de l’ADN (en cyan) (image reproduite avec la permission de Bruno Saubaméa, UMR-S 1144). |
Plus d’un siècle après l’introduction du concept de BHE par les médecins allemands Paul Ehrlich (qui a reçu en 1908 le prix Nobel de Physiologie ou Médecine, partagé avec Elie Metchnikoff pour leurs travaux sur l’immunité) et Edwin Goldman (un chirurgien disciple d’Ehrlich), de très nombreux travaux ont contribué à décrire l’organisation anatomique, cellulaire et fonctionnelle de cette barrière. Grâce aux progrès récents [4] réalisés dans le domaine de la biologie cellulaire et moléculaire, de la biochimie et de la neurocinétique des échanges entre le sang et le cerveau, les principaux mécanismes de transports au travers de la BHE ont été élucidés (Figure 2). Ces mécanismes jouent un rôle majeur dans la régulation et le maintien de l’homéostasie cérébrale pour de nombreuses molécules endogènes (sucres, acides aminés, métabolites, électrolytes, nucléosides, métaux, vitamines, hormones, etc.), mais également dans la protection du cerveau vis-à-vis de de composants présents dans la circulation générale et potentiellement neurotoxiques, qu’ils soient endogènes ou xénobiotiques. La particularité des cellules constituant la BHE par rapport aux cellules endothéliales périphériques réside dans la présence de jonctions intercellulaires serrées qui limitent très efficacement le passage paracellulaire (entre les cellules) des composés, y compris ceux de très faible masse moléculaire (de quelques dizaines de daltons). Le franchissement de la BHE par les médicaments ne peut donc être réalisé qu’au travers des cellules endothéliales (par la voie transcellulaire) et dépend de leurs propriétés physico-chimiques et/ou de leur affinité pour les systèmes de transport qui sont exprimés sur la face luminale (face sanguine) ou abluminale (face cérébrale) des cellules endothéliales constituant la BHE. Les transporteurs actifs de médicaments de type solute carrier (SLC) et ATP-binding cassette (ABC) jouent un rôle prépondérant dans ces échanges membranaires de molécules de faible masse moléculaire (moins de 4 000 Da), les transporteurs SLC permettant principalement l’influx de composés du sang vers le cerveau et les transporteurs ABC s’opposant à leur entrée dans le cerveau [5].
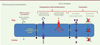 |
Figure 2. Mécanismes généraux du passage des composés à travers la barrière hémato-encéphalique. |
La transcytose est un mode particulier de passage à travers des membranes biologiques de molécules de plus haute masse moléculaire (peptides, polypeptides, protéines). Elle repose essentiellement sur deux mécanismes: la transcytose par adsorption et la transcytose dépendant de récepteurs (Figure 2). Dans le cas de la transcytose par adsorption (TMA), la formation des vésicules intracellulaires ayant pour origine la membrane plasmique résulte de phénomènes d’attractions électrostatiques. Les protéines cationiques pénètrent ainsi dans le cerveau via ce mécanisme qui est déclenché par une interaction électrostatique entre des peptides ou des protéines polycationiques et des microdomaines anioniques exprimés à la membrane des cellules endothéliales du cerveau, telles que des glycoprotéines contenant des résidus d’acide sialique à la membrane plasmique luminale, et des héparane sulfates du côté abluminal. Le(s) mécanisme(s) exact(s) d’interaction des protéines basiques avec la membrane, leur translocation dans les cellules, et leur échappement endosomal dans le cytosol permettant une sortie ultérieure, restent peu documentés. Les molécules internalisées se déplacent vers le pôle opposé de la cellule et traversent à nouveau la membrane. Ce trafic dans le cytoplasme peut entraîner l’accumulation des protéines endocytées dans le compartiment endosomal et/ou lysosomal où elles seront recyclées ou dégradées. La TMA nécessite, en général, une interaction avec des composés exprimés du côté luminal des cellules. Le mécanisme exact n’est cependant pas complètement compris et savoir si des récepteurs spécifiques sont impliqués ou si une adsorption dépendant d’énergie est nécessaire reste à déterminer.
La transcytose impliquant des récepteurs (ou TMR) nécessite, quant à elle, l’interaction de la macromolécule avec un récepteur exprimé à la surface luminale des cellules endothéliales de la barrière. Ce mécanisme permet par exemple à des molécules de très haute masse moléculaire, comme les lipoprotéines, l’insuline, la transferrine et la leptine, de franchir la membrane. Ces mécanismes de transcytose, TMA ou TMR, sont particulièrement étudiés afin d’améliorer le passage de médicaments biologiques de haute masse moléculaire au travers de la BHE.
Stratégies d’optimisation de la pénétration cérébrale d’anticorps thérapeutiques
L’efficacité d’un anticorps thérapeutique dépend fortement de ses propriétés pharmacocinétiques. Ces propriétés reposent sur plusieurs caractéristiques, dont leur activité de liaison à leur cible, leur charge, leur degré de glycosylation et leur capacité de recyclage. Les anticorps sont des protéines de haute masse moléculaire, d’environ 150 kDa. Ils sont polaires et se déplacent lentement entre les cellules endothéliales vasculaires périphériques du sang vers les espaces interstitiels des tissus. Les concentrations les plus élevées d’anticorps sont ainsi observées dans des tissus très richement perfusés et présentant un réseau vasculaire lâche, tels que la moelle osseuse, la rate ou le foie. La pénétration des anticorps est beaucoup plus limitée dans des tissus comme le cerveau, protégé par la BHE, et les tumeurs dont les réseaux de vascularisation sont très peu en contact avec la circulation lymphatique. Une des stratégies principales pour accroître la concentration d’anticorps thérapeutiques dans le tissu tumoral consiste à cibler un ou plusieurs antigènes spécifiquement surexprimés par les tumeurs [4]. Nous exposerons ici essentiellement les résultats obtenus et les essais en cours visant à faciliter le passage des anticorps au travers de la BHE et accroître ainsi l’exposition cérébrale à ces anticorps. Nous nous limiterons aux anticorps administrés par voie systémique (ce qui représente la majorité des cas). Nous ne traiterons donc pas des administrations par les autres voies (intrathécale, intracérébrale et intranasale notamment) ni les rares cas de formulations nanoparticulaires ou d’administrations facilitées par des ultrasons.
Alors que les concentrations d’immunoglobulines (IgG) dans les organes richement perfusés sont généralement comprises entre 10 et 50 % de leur taux plasmatique [6], celles dans le cerveau ne représentent, en moyenne, que de 0,01 % à 0,4 % du taux plasmatique [1]. L’une des façons d’augmenter l’exposition cérébrale d’un anticorps administré par voie systémique est donc de lui adjoindre un « module » spécifique qui lui permettra de traverser la BHE par transcytose.
Plusieurs stratégies ont été développées au cours des dernières décennies pour administrer des protéines au cerveau. L’une d’elles repose sur l’ajout de charges positives aux protéines ciblant la TMA. L’équipe de William Pardridge, physiologiste américain de l’université de Californie à Los Angeles, avait en effet montré, dès 1987, que les IgG cationisées par l’ajout de résidus polyamines, présentaient une meilleure distribution cérébrale que des IgG natives [7]. Toutefois, l’affinité de ces anticorps cationisés est souvent réduite, ce qui peut limiter leurs activités biologiques. Le cytosol des cellules, qui est un milieu réducteur, peut également altérer la structure des IgG en réduisant les ponts disulfures intra-domaines, en particulier celui du domaine variable, entraînant une modification conformationnelle avec, in fine, une perte de reconnaissance de l’antigène par l’anticorps. La TMA n’est par ailleurs pas spécifique d’un type de tissu ou d’une cellule. Des effets secondaires touchant différentes cibles peuvent donc apparaître en cas d’utilisation thérapeutique. Enfin, ces mécanismes d’endocytose qui dépendent des charges des molécules sont souvent considérés comme étant toxiques.
D’autres stratégies ont été élaborées afin de cibler les mécanismes de TMR. Elles consistent à utiliser des modules transporteurs qui sont inspirés de ceux présents au niveau de la BHE elle-même et des mécanismes qui lui permettent d’importer les protéines endogènes qui lui sont indispensables. Les transporteurs les plus étudiés aujourd’hui sont ceux de l’insuline et de la transferrine. Des travaux pionniers, conduits par William Pardridge [8], ont montré l’intérêt de ces transporteurs pour véhiculer des protéines dans le cerveau et ont inspiré une variété de formats d’anticorps multi-spécifiques qui ont permis d’augmenter leur exposition cérébrale, parfois d’un facteur important. Ces nouveaux formats sont, par exemple, des anticorps contre les récepteurs de l’insuline ou de la transferrine auxquels est fusionnée une protéine thérapeutique qui peut-être une enzyme, un facteur de croissance, un anticorps ou un de ses fragments (comme par exemple un scFv [single chain fragment variable] ou un Fab).
Quelques constructions appartenant à cette catégorie (Figure 3A) sont en développement clinique, comme des protéines de fusion constituées de l’enzyme lysosomale iduronate sulfatase fusionnée avec un anticorps contre le récepteur de l’insuline [9] (Figure 3A, -1-) ou avec un anticorps contre le récepteur de la transferrine [10] (Figure 3A, -2-), avec lesquelles des études cliniques de phase II sont en cours pour traiter la mucopolysaccharidose2. Plusieurs protéines de fusion de ce type ont été construites avec des anticorps anti-bêta-amyloïde soit sous forme de fragments ScFv ( Figure 3A, -3-) [11] ou de fragments F(ab’)2 de ces anticorps [12] (Figure 3A, -4-). Elles ont montré un intérêt dans des modèles murins ou en imagerie de la maladie d’Alzheimer [13]. Des protéines de fusion avec d’autres protéines thérapeutiques ont été décrites, comme celle fusionnée avec l’érythropoiëtine [14], ou avec une protéine inhibitrice du TNF-a (tumor necrosis factor alpha) [15] ou avec des enzymes lysosomales [16].
 |
Figure 3. Nouveaux formats d’anticorps thérapeutiques à pénétration cérébrale renforcée. BACE1: beta-secretase 1. |
Une deuxième catégorie de constructions (Figure 3B) consiste en des anticorps thérapeutiques dans lesquels un paratope3 spécifique a été incorporé. Ce paratope peut reconnaître le récepteur de la transferrine, soit sous forme de bispécifiques bivalents (Figure 3B, -6-)comme les DVD (dual variable domain) [17] ou les TBTI (tetravalent bispecific tandem IgG) [18] ou monovalents (Figure 3B, -7-) [19], ou résultant de la fusion d’un domaine scFv (Figure 3B, -8-) [20]. Là encore, des anticorps se sont révélés prometteurs dans la maladie d’Alzheimer [un anticorps contre la protéine beta-amyloïde (Figure 3B, -6,8-) et un anticorps contre l’enzyme BACE1 (beta-secretase 1) (Figure 3B, -7-) qui est responsable des coupures de la protéine à l’origine des fragments toxiques].
La pénétration des anticorps dans les tissus repose sur leur affinité pour leurs cibles. Ce processus, qui a été décrit pour la distribution des anticorps au sein des tumeurs [21], a été également observé pour l’exposition d’anticorps dirigés contre le récepteur de la transferrine dans le cerveau. Un laboratoire américain a ainsi montré que des mutations touchant le paratope d’anticorps spécifiques de ce récepteur, affectant leur affinité pour leur cible, pouvaient augmenter, dans certaines conditions, leur exposition dans le parenchyme cérébral [18]. Un grand groupe pharmaceutique européen a également décrit le fait que certains formats d’anticorps monovalents, spécifiques du récepteur de la transferrine (Figure 3B, -7-), qui ont une avidité réduite pour leur cible, présentent des expositions cérébrales augmentées [20]. Le potentiel de ces avancées permettant d’accroître les expositions cérébrales d’anticorps dirigés contre des récepteurs exprimés par la BHE est très prometteur dans toutes les indications concernant le système nerveux central, que ce soient les maladies neurodégénératives ou les désordres centraux liés à certaines maladies génétiques, ou le glioblastome, les lymphomes cérébraux et les métastases cérébrales. Quelques formats sont d’ores et déjà en développement. Ils ont fait la preuve de leur innocuité. Néanmoins, pour certains, comme les anticorps contre les récepteurs de l’insuline ou de la transferrine, un ajustement très fin de leurs propriétés reste nécessaire afin d’éviter, pour les uns, des problèmes de toxicité liés à leurs effets sur la glycémie [22] ou des désordres pancréatiques [23], ou, pour les autres, des désordres sanguins ou médullaires [24]. L’avenir nous dira si ce potentiel livre toutes ses promesses.
De nouveaux formats d’anticorps ont fait récemment leur apparition. Ils sont issus d’immunoglobulines d’autres espèces animales que l’homme. Les camélidés (chameaux, dromadaires, lamas, alpagas, etc.) ont la particularité de produire des anticorps conventionnels possédant des chaînes lourdes et légères mais aussi deux sous-classes d’IgG formées uniquement de dimères de chaînes lourdes [25]. Le paratope de ces anticorps à domaine unique est constitué d’un seul domaine VH, appelé VHH ou nanobodyTM. Ces anticorps homodimériques dont la chaîne lourde se caractérise par l’absence de domaine CH1, révèlent ainsi que le répertoire anticorps ne repose pas uniquement sur une combinatoire VH-VL. Des anticorps à domaine unique ont aussi été trouvés dans les poissons cartilagineux comme le requin. Leurs fragments (VNAR) présentent le même type de propriétés que les VHH [26]. Un grand nombre d’anticorps à domaine unique ayant une affinité élevée vis-à-vis d’un antigène ont déjà été isolés par la technique du phage display4 ou par ribosome display5. En raison de leur pouvoir de diffusion élevé lié à leur petite taille (15 kDa), de leur sélectivité et de l’absence de réactions immunologiques lorsqu’ils sont injectés dans des animaux d’autres espèces ou chez l’homme, ces VHH combinent les avantages des anticorps conventionnels à ceux des petites molécules [27]. Ils sont aussi naturellement très stables: ils supportent des températures élevées et la réduction de leur pont disulfure intra-domaine n’entraîne pas de perte de reconnaissance de l’antigène qu’ils ciblent. Cette dernière propriété permet ainsi d’utiliser ces VHH, après leur transfection à l’intérieur d’une cellule, comme marqueur intracellulaire [28]. Environ 2/3 des VHH ont un point isoélectrique basique. Ils sont donc naturellement cationisés ce qui leur confère la capacité de traverser facilement la BHE in vivo. Lafaye et al. ont en effet montré que des VHH ayant un point isoélectrique basique et dirigés contre la protéine fibrillaire gliale acide (glial fibrillary acidic protein, ou GFAP), un marqueur spécifique des astrocytes, étaient capables, après leur injection intra-carotidienne chez la souris, de se lier à cette protéine dans le cytoplasme des astrocytes [29]. Cette capacité des VHH de traverser la barrière a été confirmée récemment avec des VHH dirigés contre les deux principales lésions de la maladie d’Alzheimer: les plaques amyloïdes intra-cérébrales et les dégénérescences neurofibrillaires (DNF) intra-neuronales. Ces VHH marqués par un fluorochrome vert ont en effet été testés in vivo dans deux modèles murins de maladie d’Alzheimer. Après injection par voie intraveineuse, les VHH traversent la BHE et interagissent avec les plaques amyloïdes et les DNF [30] ; une étude pharmacocinétique utilisant le VHH dirigé contre les plaques amyloides montre que, deux heures après l’injection intraveineuse, 0,5 % de la dose injectée est retrouvé dans le cerveau. Le marquage intracérébral que permettent ces 3 VHH repose sur un ensemble de propriétés que les anticorps conventionnels n’ont pas: (1) la capacité de traverser la BHE et d’entrer dans les cellules, car ils sont cationisés ; (2) une petite taille permettant leur diffusion dans le tissu cérébral ; (3) la reconnaissance spécifique d’un antigène intracellulaire grâce à leur très grande stabilité ; et, enfin, (4) leur capacité à transporter des molécules pour le diagnostic ou la thérapie.
Conclusion
Le cerveau est probablement l’ultime frontière pour les anticorps, aussi bien pour le diagnostic que pour la thérapie. Pour qu’une approche cérébrale de la thérapie par anticorps soit efficace, la définition des cibles est cruciale. Ceci implique une meilleure compréhension des mécanismes physiopathologiques impliqués dans le développement des pathologies touchant le cerveau. L’étude des mécanismes permettant le passage des anticorps au travers de la BHE nécessite également d’explorer de nouvelles pistes, comme par exemple l’ouverture transitoire de la BHE en utilisant des ultrasons, ce qui pourrait faciliter l’entrée de ces anticorps au sein du cerveau [31, 32].
Liens d’intérêt
Les auteurs déclarent n’avoir aucun lien d’intérêt concernant les données publiées dans cet article.
Les péricytes sont des cellules localisées au niveau de la lame basale de l’endothélium des capillaires. Ils l’entourent par de longs prolongements cytoplasmiques.
Maladie rare du stockage lysosomal due à des mutations alléliques du gène codant l’enzyme.
Partie de l’anticorps reconnaissant l’épitope porté par l’antigène.
Cette technique permet l’expression de peptides ou de protéines exogènes à la surface des particules de phages, afin de sélectionner et d’amplifier un polypeptide capable d’interagir avec la molécule cible choisie.
Le ribosome display est une technique de sélection des ligands (ici un anticorps) où le lien entre le phénotype et le génotype est réalisé par un complexe ternaire formé entre la protéine traduite, le ribosome et l’ARN messager correspondant.
Références
- Pepinsky RB. Shao Z. Ji B, et al. Exposure levels of anti-LINGO-1 Li81 antibody in the central nervous system and dose-efficacy relationships in rat spinal cord remyelination models after systemic administration. J Pharmacol Exp Ther 2011 ; 339: 519–529. [Google Scholar]
- Strazielle N. Ghersi-Egea JF. Physiology of blood-brain interfaces in relation to brain disposition of small compounds and macromolecules. Mol Pharm 2013 ; 10: 1473–1491. [CrossRef] [PubMed] [Google Scholar]
- Abbott NJ. Blood-brain barrier structure and function and the challenges for CNS drug delivery. J Inherit Metab Dis 2013 ; 36: 437–449. [CrossRef] [PubMed] [Google Scholar]
- Husain B. Ellerman D. Expanding the boundaries of biotherapeutics with bispecific antibodies. BioDrugs 2018 ; 32: 441–464. [CrossRef] [PubMed] [Google Scholar]
- Chaves C. Shawahna R. Jacob A, et al. Human ABC transporters at blood-CNS interfaces as determinants of CNS drug penetration. Curr Pharm Des 2014 ; 20: 1450–1462. [CrossRef] [PubMed] [Google Scholar]
- Lobo ED. Hansen RJ. Balthasar JP. Antibody pharmacokinetics and pharmacodynamics. J Pharm Sci 2004 ; 93: 2645–2668. [CrossRef] [PubMed] [Google Scholar]
- Kumagai AK. Eisenberg JB. Pardridge WM. Absorptive-mediated endocytosis of cationized albumin and a beta-endorphin-cationized albumin chimeric peptide by isolated brain capillaries. Model system of blood-brain barrier transport. J Biol Chem 1987 ; 262: 15214–15219. [PubMed] [Google Scholar]
- Pardridge WM. Delivery of biologics across the blood-brain barrier with molecular trojan horse technology. BioDrugs 2017 ; 31: 503–519. [CrossRef] [PubMed] [Google Scholar]
- Pardridge WM. Boado RJ. Giugliani R. Schmidt M. Plasma pharmacokinetics of valanafusp alpha, a human insulin receptor antibody-iduronidase fusion protein, in patients with mucopolysaccharidosis type I. BioDrugs 2018 ; 32: 169–176. [CrossRef] [PubMed] [Google Scholar]
- Sonoda H. Morimoto H. Yoden E, et al. A Blood-brain-barrier-penetrating anti-human transferrin receptor antibody fusion protein for neuronopathic mucopolysaccharidosis II. Mol Ther 2018 ; 26: 1366–1374. [CrossRef] [PubMed] [Google Scholar]
- Sumbria RK. Zhou QH. Hui EK, et al. Pharmacokinetics and brain uptake of an IgG-TNF decoy receptor fusion protein following intravenous, intraperitoneal, and subcutaneous administration in mice. Mol Pharm 2013 ; 10: 1425–1431. [CrossRef] [PubMed] [Google Scholar]
- Sehlin D. Fang XT. Meier SR, et al. Pharmacokinetics, biodistribution and brain retention of a bispecific antibody-based PET radioligand for imaging of amyloid-beta. Sci Rep 2017 ; 7: 17254. [CrossRef] [PubMed] [Google Scholar]
- Syvanen S. Fang XT. Hultqvist G, et al. A bispecific Tribody PET radioligand for visualization of amyloid-beta protofibrils - a new concept for neuroimaging. NeuroImage 2017 ; 148: 55–63. [CrossRef] [PubMed] [Google Scholar]
- Chang R. Al Maghribi A. Vanderpoel V, et al. Brain penetrating bifunctional erythropoietin-transferrin receptor antibody fusion protein for Alzheimer’s disease. Mol Pharm 2018 ; 15: 4963–4973. [CrossRef] [PubMed] [Google Scholar]
- Chang R. Knox J. Chang J, et al. Blood-brain barrier penetrating biologic TNF-alpha inhibitor for Alzheimer’s disease. Mol Pharm 2017 ; 14: 2340–2349. [CrossRef] [PubMed] [Google Scholar]
- Boado RJ. Lu JZ. Hui EK, et al. Insulin receptor antibody-alpha-N-acetylglucosaminidase fusion protein penetrates the primate blood-brain barrier and reduces glycosoaminoglycans in Sanfilippo type B fibroblasts. Mol Pharm 2016 ; 13: 1385–1392. [CrossRef] [PubMed] [Google Scholar]
- Karaoglu Hanzatian D, Schwartz A, Gizatullin F, et al. Brain uptake of multivalent and multi-specific DVD-Ig proteins after systemic administration. mAbs 2018; 10: 765–77. [CrossRef] [PubMed] [Google Scholar]
- TM. Do, I. Arnould, J. Beninga, et al. Brain exposure and therapeutic efficacy of multivalent bispecific anti-TfRC antibodies. Abstracts from the 22nd International Symposium on signal transduction at the blood-brain barriers. Fluids Barriers CNS 2019; 16 (suppl 2): 29. [CrossRef] [PubMed] [Google Scholar]
- Yu YJ, Zhang Y, Kenrick M, et al. Boosting brain uptake of a therapeutic antibody by reducing its affinity for a transcytosis target. Sci Transl Med 2011; 3: 84ra44. [PubMed] [Google Scholar]
- Niewoehner J. Bohrmann B. Collin L, et al. Increased brain penetration and potency of a therapeutic antibody using a monovalent molecular shuttle. Neuron 2014 ; 81: 49–60. [CrossRef] [PubMed] [Google Scholar]
- Adams GP. Schier R. McCall AM, et al. High affinity restricts the localization and tumor penetration of single-chain fv antibody molecules. Cancer Res 2001 ; 61: 4750–4755. [Google Scholar]
- Boado RJ. Hui EK. Lu JZ. Pardridge WM. Glycemic control and chronic dosing of rhesus monkeys with a fusion protein of iduronidase and a monoclonal antibody against the human insulin receptor. Drug Metab Dispos 2012 ; 40: 2021–2025. [CrossRef] [PubMed] [Google Scholar]
- Ohshima-Hosoyama S. Simmons HA. Goecks N, et al. A monoclonal antibody-GDNF fusion protein is not neuroprotective and is associated with proliferative pancreatic lesions in parkinsonian monkeys. PLoS One 2012 ; 7: e39036. [CrossRef] [PubMed] [Google Scholar]
- Pardridge WM. Boado RJ. Patrick DJ, et al. Blood-brain barrier transport, plasma pharmacokinetics, and neuropathology following chronic treatment of the rhesus monkey with a brain penetrating humanized monoclonal antibody against the human transferrin receptor. Mol Pharm 2018 ; 15: 5207–5216. [CrossRef] [PubMed] [Google Scholar]
- Hamers-Casterman C. Atarhouch T. Muyldermans S, et al. Naturally occurring antibodies devoid of light chains. Nature 1993 ; 363: 446–448. [Google Scholar]
- Feng M, Bian H, Wu X, et al. Construction and next-generation sequencing analysis of a large phage-displayed VNAR single-domain antibody library from six naïve nurse sharks”. Antibody Therapeutics 2019; 2 h 1–11. [Google Scholar]
- Nguyen VK. Desmyter A. Muyldermans S. Functional heavy-chain antibodies in Camelidae. Adv Immunol 2001 ; 79: 261–296. [CrossRef] [PubMed] [Google Scholar]
- Traenkle B. Rothbauer U. Under the microscope: Single-domain antibodies for live-cell imaging and super-resolution microscopy. Front Immunol 2017 ; 8: 1030. [CrossRef] [PubMed] [Google Scholar]
- Li T. Bourgeois JP. Celli S, et al. Cell-penetrating anti-GFAP VHH and corresponding fluorescent fusion protein VHH-GFP spontaneously cross the blood-brain barrier and specifically recognize astrocytes: application to brain imaging. FASEB J 2012 ; 26: 3969–3979. [CrossRef] [PubMed] [Google Scholar]
- Li T. Vandesquille M. Koukouli F, et al. Camelid single-domain antibodies: a versatile tool for in vivo imaging of extracellular and intracellular brain targets. J Control Release 2016 ; 243: 1–10. [CrossRef] [PubMed] [Google Scholar]
- Carpentier A, Canney M, Vignot A, et al. Clinical trial of blood-brain barrier disruption by pulsed ultrasound. Sci Transl Med 2016; 8: 343re2. [PubMed] [Google Scholar]
- Santin MD. Debeir T. Bridal SL, et al. Fast in vivo imaging of amyloid plaques using mu-MRI Gd-staining combined with ultrasound-induced blood-brain barrier opening. NeuroImage 2013 ; 79: 288–294. [CrossRef] [PubMed] [Google Scholar]
Liste des figures
|
Figure 1. Structure histologique de la barrière hémato-encéphalique (BHE). A. Schéma d’un capillaire cérébral en coupe transversale montrant les différentes structures cellulaires composant la BHE. B. Visualisation par microscopie confocale d’un vaisseau cérébral dans le cortex chez le rat. Les astrocytes sont révélés par un immunomarquage de la GFAP (glial fibrillary acidic protein) (en jaune) et la membrane luminale des cellules endothéliales par un immunomarquage de la P-glycoprotéine (en magenta). Les noyaux cellulaires (cellules endothéliales, cellules murales, neurones, cellules gliales) sont marqués par un intercalant de l’ADN (en cyan) (image reproduite avec la permission de Bruno Saubaméa, UMR-S 1144). |
| Dans le texte | |
|
Figure 2. Mécanismes généraux du passage des composés à travers la barrière hémato-encéphalique. |
| Dans le texte | |
|
Figure 3. Nouveaux formats d’anticorps thérapeutiques à pénétration cérébrale renforcée. BACE1: beta-secretase 1. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.