")
")
| Issue |
Med Sci (Paris)
Volume 30, Number 11, Novembre 2014
Cils primaires et ciliopathies
|
|
|---|---|---|
| Page(s) | 955 - 961 | |
| Section | Cils primaires et ciliopathies | |
| DOI | https://doi.org/10.1051/medsci/20143011008 | |
| Published online | 10 novembre 2014 | |
Élongation de l’axonème et dynamique du transport intraflagellaire
Elongation of the axoneme and dynamics of intraflagellar transport
Unité de biologie cellulaire des trypanosomes, Institut Pasteur et CNRS, 25, rue du docteur Roux, 75015
Paris, France
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
**
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Résumé
Les cils et flagelles sont des organites essentiels chez la plupart des eucaryotes, y compris chez l’humain. Dans cette revue, nous abordons le mode d’assemblage de ces organites complexes, qui fait intervenir un processus dynamique dénommé transport intraflagellaire. Le transport intraflagellaire apporte les éléments structuraux au bout distal du cil où a lieu l’assemblage, permettant ainsi la croissance de l’organite. Nous exposons ensuite trois modèles différents de contrôle de la longueur du cil avant de discuter de leur altération lors des ciliopathies, maladies génétiques associées aux défauts ciliaires.
Abstract
Cilia and flagella are essential organelles in most eukaryotes including human beings. In this review, we will discuss the mode of assembly of these complex organelles that depends on a dynamic process called intraflagellar transport or IFT. IFT delivers structural elements at the distal end of the cilium where assembly takes place, thereby allowing the growth of the organelle. We next discuss the different models for control of cilium length and their alterations in ciliopathies, genetic diseases associated to ciliary defects.
Cet article fait partie du numéro thématique de médecine/sciences intitulé « Cils primaires et ciliopathies » (m/s n° 11, vol. 30, novembre 2014).
© 2014 médecine/sciences – Inserm
Cils et flagelles : structure et composition
Ces pieds incroyablement minces ou petites jambes qui ont été déplacés avec agilité » ; c’est par ces mots que Antoni van Leeuwenhoek décrivit pour la première fois les cils et les flagelles en 1675. Les termes « cils » et « flagelles » ont été introduits au xviiie et xixe siècles pour décrire ces organites présents à la surface de nombreuses cellules eucaryotes. L’utilisation d’une dénomination différente pour le cil et le flagelle est historique et, comme nous allons le voir, l’organisation structurale de ces deux organites est équivalente. Nous emploierons dès lors les deux termes de manière interchangeable. Il s’agit de structures cylindriques constituées d’un axonème composé de neuf doublets de microtubules entourés d’une membrane (Figures 1, 2). Deux configurations principales peuvent être rencontrées, « 9+2 » et « 9+0 », selon que les neuf doublets de microtubules périphériques entourent ou non une paire centrale de singulets de microtubules.
-
Dans la structure « 9+2 », des moteurs moléculaires appartenant à la famille des dynéines sont ancrés sous forme de bras sur les microtubules périphériques (Figure 2E). La paire centrale de microtubules, les ponts radiaires et le DRC (dynein regulatory complex)/nexine régulent l’activité des moteurs de dynéines qui génèrent la force nécessaire pour le battement. Cette configuration est typique de la plupart des cils mobiles. Par exemple, les cellules épithéliales qui tapissent l’épendyme des mammifères présentent une cinquantaine de cils mobiles assurant la circulation du liquide céphalorachidien (Figure 1A).
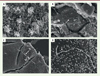
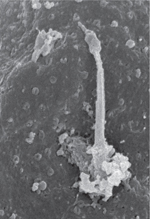
Figure 1. Diversité du nombre, de la longueur et de la disposition des cils. A. Épithélium tapissant les ventricules du cerveau de la souris. B. Épithélium du tube collecteur du rein de souris visualisé après cryofracture mécanique. C. Cil primaire à la surface de cellules IMCD (inner medullary collecting duct) et ( D ) de cellules MDCK (Mardin Darby canine kidney). Toutes les images ont été prises au microscope électronique à balayage et ont été modifiées à partir de Vincensini et al. [40]. Barres d’échelle : 1 µm. Les cils sont indiqués par des flèches.

Figure 2. Ultrastructure du flagelle de Trypanosoma brucei. A. Coupe longitudinale de la base du flagelle. Le corps basal immature n’est pas visible sur cette section. Les mentions B-E dans le panneau A indiquent les sections transversales à travers le flagelle dont les champs sont grossis dans la colonne de droite de la figure. B. Corps basal. C. Le début de la zone de transition avec les fibres de transition dont l’extrémité est indiquée par des flèches. D. La portion centrale de la zone de transition. Les têtes de flèches indiquent la collerette. E. Axonème dans la lumière de la poche flagellaire. Du matériel dense aux électrons est fréquemment observé entre les doublets périphériques et la membrane. F. Représentation schématique de la base du flagelle indiquant le corps basal (9 triplets de microtubules), la zone de transition (9 doublets de microtubules) et l’axonème (9 doublets de microtubules représentés en bleu foncé et une paire de singulets représentés en bleu clair). FPC : collier de la poche flagellaire ; K : kinétoplaste ; G : appareil de Golgi. Barres d’échelle : 100 nm. Les photographies sont extraites de [41].
-
En revanche, dans la structure « 9+0 », la paire centrale de microtubules, les moteurs de dynéines et tous les autres composants nécessaires à la fonction de battement sont absents. Cette configuration est celle de la majorité des cils sensoriels comme ceux qui sont rencontrés dans le rein (Figure 1B), ou du cil primaire présent à la surface de très nombreuses cellules du corps humain (Figure 1C, D). Bien que le cil primaire ait été découvert à la fin du xixe siècle, son rôle a été longtemps ignoré, jusqu’à la découverte de sa contribution au bon fonctionnement du rein [1]. Le cil primaire est immobile, mais il contient un nombre important d’éléments sensoriels ; il est essentiel au développement de la plupart des organes.
Les cils et les flagelles ont été conservés chez la majorité des eucaryotes au cours de l’évolution, avec de rares exceptions [2]. Toutefois, le nombre, la longueur et la position des cils et des flagelles varient fortement d’un organisme à un autre, et même d’un type cellulaire à l’autre chez les organismes multicellulaires. Des analyses protéomiques de cils ou flagelles purifiés à partir de différentes espèces ont mis en évidence la sophistication de ces organites, qui contiennent plus de 500 protéines dont la moitié sont conservées [3–5]. Dans la majorité des cas, il s’agit de protéines essentielles à la structure, à la construction ou au mouvement de l’organite.
Le cil, un compartiment bien individualisé
Les cils constituent un compartiment séparé du reste du corps cellulaire, non pas par une membrane, mais par des éléments structuraux uniques (Figure 2). Le cil est constitué de trois compartiments : le corps basal (Figure 2B), la zone de transition (Figure 2C, D) et l’axonème (Figure 2E). Le corps basal peut être considéré comme un centriole modifié ; il est composé de neuf triplets de microtubules (A, B, C) [6] (Figure 2B). Deux d’entre eux (A et B) se poursuivent pour constituer l’armature de la zone de transition (Figure 2D) et de l’axonème (Figure 2E). Le corps basal est ancré à la membrane plasmique par des fibres de transition qui délimitent le compartiment flagellaire et le séparent du reste de la cellule (Figure 2C). Les fibres de transition permettent d’attacher les doublets de microtubules à la membrane. Elles pourraient également servir de site d’amarrage aux protéines entrant dans le flagelle [7]. La zone de transition est comprise entre le corps basal et l’axonème. C’est à son niveau que la membrane flagellaire commence. Des projections denses aux électrons, en forme de « Y », connectent chaque doublet de microtubules à la membrane ciliaire. Elles semblent associées à un « collier de perles » qui entoure la membrane du cil seulement à cet endroit [8]. L’ensemble est responsable d’une séparation physique du flagelle avec le cytoplasme. Ce rôle de filtre moléculaire opère aussi bien pour les protéines de la matrice ciliaire que pour celles de la membrane [9].
Construction du flagelle et transport intraflagellaire
La construction des cils et flagelles nécessite la production et l’assemblage correct d’un grand nombre de protéines précisément organisées à la fois dans le temps et dans l’espace. Dans cette revue, nous décrivons principalement les résultats obtenus chez Chlamydomonas reinhardtii et Trypanosoma brucei, deux modèles d’étude qui ont fourni des informations essentielles pour la compréhension du mode de formation des cils et flagelles. Le cil étant dépourvu de ribosomes, les protéines qui le composent doivent être synthétisées dans le cytoplasme. Au cours de l’élongation de l’axonème, l’incorporation de protéines a lieu à l’extrémité distale de l’organite [10]. Un système de transport appelé transport intraflagellaire (IFT, intraflagellar transport) a été découvert pour la première fois chez l’algue verte, Chlamydomonas, par microscopie à contraste interférentiel1. Il s’agit d’un mouvement de particules (ou trains) disposées entre la membrane et les doublets périphériques et qui se déplacent, soit de la base vers l’extrémité distale du flagelle (transport antérograde), soit dans le sens inverse (transport rétrograde) [11]. Chez Chlamydomonas, les trains de particules ont une longueur d’environ 700 nm pour 80 nm de largeur et 25 nm de hauteur (transport antérograde), tandis que les trains retour (transport rétrograde) ont une longueur avoisinant les 250 nm pour 50 nm de largeur et 20 nm de hauteur [12].
Le transport antérograde est conduit par la kinésine II, qui entraîne les trains de particules de la base du flagelle à son extrémité distale où est localisée l’extrémité (+) des microtubules (site de polymérisation/dépolymérisation) [13]. Un complexe dynéine spécifique au transport intraflagellaire est responsable du transport rétrograde et se charge de ramener les trains de particules à la base du flagelle où se situe le bout (-) des microtubules [14, 15] (Figure 3).
 |
Figure 3. Modèle du transport intraflagellaire. (1) Les particules IFT, les moteurs et les précurseurs axonémaux sont recrutés à la base du flagelle. (2) L’assemblage des particules IFT et le chargement des précurseurs axonémaux auraient lieu au niveau des fibres de transition (représentées en noir). (3) La kinésine II et le complexe IFT-B sont indispensables au transport antérograde en direction de la pointe du flagelle. La dynéine est transportée sous une forme inactive. (4) Les particules IFT sont remaniées à l’extrémité distale du flagelle. (5) Les particules retournent à la base du flagelle par l’intermédiaire de la dynéine, responsable du transport rétrograde, pour y être recyclées [24]. La kinésine est alors inactivée et revient vers la base soit sous forme de cargo [15], soit par diffusion [30]. KAP : kinesin accessory protein ; DIC5 : chaîne intermédiaire de dynéine ; DLIC1 : chaîne légère intermédiaire de dynéine ; DHC2.1 : chaîne lourde de dynéine 2. |
Les particules constituant ce système de transport ont été purifiées par centrifugation différentielle à partir de flagelles isolés de Chlamydomonas, permettant l’identification de la plupart des protéines les composants [16]. Deux sous-complexes ont été identifiés : le complexe IFT-A, constitué de six protéines, et le complexe IFT-B, composé d’au minimum onze protéines [16, 17]. Des études in vitro ont permis de révéler l’organisation moléculaire de ces deux complexes [18]. Les protéines des complexes IFT sont localisées le long de l’axonème, mais une proportion significative d’entre elles sont concentrées à la base du flagelle [16, 19–22]. L’expression de ces protéines fusionnées à la GFP (green fluorescent protein) permet de révéler leur mouvement au sein des cils et flagelles [19, 21, 23]. Les trains de particules, et donc de protéines, sont transportés par la kinésine II jusqu’à l’extrémité distale, puis repartent rapidement dans le sens opposé par l’intermédiaire de la dynéine. Que se passe-t-il à l’extrémité distale du flagelle ? Comme les trains antérogrades mesurent environ 700 nm et que les trains rétrogrades sont trois fois plus petits, on peut imaginer que les premiers sont alors scindés en trois parties. Cette hypothèse est confirmée par l’analyse de la fréquence des trains chez Trypanosoma brucei, qui révèle qu’il existe un train antérograde pour trois trains rétrogrades [24]. Ce processus de conversion prend au maximum trois à quatre secondes. Une fois revenus à la base du flagelle, les trains sont réutilisés pour la construction de nouveaux trains antérogrades. L’inhibition de ce transport intraflagellaire empêche l’assemblage des cils et flagelles chez la plupart des organismes étudiés jusqu’à présent [13, 15, 20, 25, 26].
L’intégrité des moteurs comme des trains IFT est essentielle. On citera ainsi les études pionnières réalisées chez Chlamydomonas au moyen du mutant thermosensible fla10 (flagellar kinesin-like protein 10) qui code pour une sous-unité du moteur de la kinésine II. Quand ces cellules sont transférées à la température restrictive, le transport intraflagellaire s’arrête et les cellules sont incapables de construire un flagelle [13]. Il a dès lors été proposé que ce processus de transport intraflagellaire assure le transport de la tubuline et des autres précurseurs flagellaires à la pointe distale du flagelle. Cette hypothèse est confortée par la mise en évidence, sur les protéines IFT74 et IFT81, d’un domaine d’homologie à la calponine qui est capable de lier la tubuline in vitro. La mutation des résidus essentiels à cette liaison ne perturbe pas la formation des trains IFT, mais inhibe la construction des cils [27].
Mécanismes de contrôle de la longueur des cils
Dans la plupart des cellules, la longueur des cils et flagelles est régulée de façon précise. Plusieurs modèles ont été proposés pour en expliquer le mode de contrôle. La situation la plus simple consiste à limiter la disponibilité de protéines axonémales dans le cytoplasme. Dans ce cas, une quantité donnée de protéines est produite et entièrement utilisée pour assembler l’organite. Lorsque cette réserve est épuisée, la croissance s’arrête. Toutefois, l’existence dans le cytoplasme d’une réserve de matériel non assemblé a été démontrée chez plusieurs espèces. Deux autres modèles ont été suggérés au cours de ces dernières années : l’autorégulation et la présence d’un senseur de longueur. Le premier modèle postule que le flagelle contient l’information nécessaire à la régulation de sa longueur, et le second qu’existe un système détectant la longueur du flagelle et capable de la moduler. Dans les deux cas, l’IFT serait l’acteur principal.
Modèle 1 : régulation par l’équilibre entre assemblage et désassemblage des sous-unités de tubuline
Les cils sont des structures dynamiques ; l’extrémité (+) de l’axonème perd continuellement des sous-unités de tubuline. Ce processus est constant et ne dépend pas de la longueur du flagelle [28]. Cette perte doit être compensée par l’addition de nouvelles sous-unités, et c’est l’équilibre entre l’assemblage et le désassemblage des sous-unités de tubuline au bout distal des microtubules qui déterminerait la longueur de l’organite [28]. Le transport intraflagellaire, qui apporte de façon continue de la tubuline à l’extrémité distale de l’axonème, serait nécessaire.
-
Contrôle de la longueur par l’apport de tubuline au bout distal. Toutefois, la quantité de tubuline fournie serait dépendante de la longueur du flagelle. En effet, la quantité totale de trains de particules dans le flagelle étant constante quelle que soit la longueur de ce dernier, les trains mettront de plus en plus de temps pour atteindre l’extrémité au fur et à mesure que le flagelle s’allonge. Lorsque le flagelle est court, l’apport de tubuline est nettement supérieur à la perte, et l’organite s’allonge rapidement (Figure 4A). Plus le flagelle s’allonge, plus son taux de croissance ralentit parce que la distance à parcourir augmente, jusqu’à atteindre l’équilibre entre les taux d’assemblage et de désassemblage. Si la croissance se poursuit au-delà du point d’équilibre, la vitesse de désassemblage est plus rapide et le flagelle raccourcit pour revenir au point d’équilibre. Ce modèle se fonde sur une diminution de la fréquence du transport intraflagellaire (que reflète le nombre de trains par unité de temps) au cours de l’allongement, qui n’a pas été confirmée par des mesures du trafic ; celles-ci suggèrent plutôt une fréquence quasi invariante au cours de l’allongement (Figure 4B) [29].
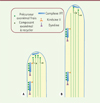
Figure 4. Modèle de l’équilibre du nombre de particules IFT. Les points verts représentent les protéines axonémales. A. Au stade précoce de formation du cil, l’apport de tubuline est supérieur à la perte et le flagelle s’allonge. B. Lorsque le cil grandit, le taux de croissance ralentit à cause de l’augmentation de la distance à parcourir jusqu’à atteindre un équilibre entre assemblage et désassemblage de la tubuline.
-
Régulation par taille des trains de particules. Une autre possibilité serait que la taille des trains2 varie au cours du temps, ce qui a été démontré chez des souches de Chlamydomonas exprimant des protéines de fusion entre la GFP et des protéines de l’IFT [30]. Aux stades précoces, les trains expriment une fluorescence importante ; ils seraient plus gros et, de ce fait, transporteraient plus de tubuline (Figure 5A). Lorsque le flagelle approche de sa longueur finale, l’intensité de fluorescence est plus faible, ce qui indique des trains plus petits, qui apportent donc moins de tubuline par unité de temps (Figure 5B). Ce modèle suppose que le taux de chargement des trains est équivalent à tous les stades [28, 30].
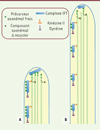
Figure 5. Modèle du contrôle de la longueur du cil par la quantité de protéines IFT disponibles dans l’organite. A. Une quantité fixe d’IFT est injectée dans le cil au stade précoce de sa formation. Les trains sont alors longs et peuvent transporter une grande quantité de précurseurs comme la tubuline dont l’apport est supérieur à la perte, permettant ainsi l’élongation du cil. B. Lorsque le cil s’allonge, les trains deviennent plus petits et apportent de moins en moins de précurseurs au bout distal, jusqu’à atteindre un équilibre dynamique.
-
Taux de chargement des trains. Récemment, le groupe de Karl Lechtreck a réussi à visualiser des précurseurs de l’axonème3 couplés à la GFP dans des cellules vivantes. Ces précurseurs se déplacent dans les flagelles simultanément avec les IFT, permettant ainsi de suivre directement leur devenir [31]. Les résultats démontrent que la quantité transportée par unité de temps diminue au cours de l’élongation du flagelle, en accord avec un modèle d’équilibre entre assemblage et désassemblage (Figure 6). Le nombre d’évènements de transport de ces précurseurs est en accord avec la taille attendue du flagelle. Dans ce cas, c’est le taux de chargement des trains – et non leur taille - qui contrôlerait la longueur du flagelle.
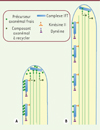
Figure 6. Modèle du contrôle de la longueur du flagelle en fonction du chargement des précurseurs axonémaux sur les particules IFT. A. Au stade précoce de construction, le chargement de précurseurs axonémaux est important et permet au cil de s’allonger rapidement. B. Le taux de chargement se réduit lorsque le flagelle s’allonge, jusqu’à atteindre l’équilibre entre assemblage et désassemblage.
Enfin, des modifications de la vitesse des trains pourraient également intervenir, assurant une livraison plus rapide ou plus lente de la tubuline, qui pourrait ainsi expliquer l’existence de flagelles plus longs ou plus courts [32, 33].
Modèle 2 : senseur de la longueur du flagelle
Par ailleurs, la longueur des cils et des flagelles pourrait être modifiée en changeant soit le taux d’assemblage, soit le taux de désassemblage. Ces taux pourraient être régulés par un senseur de la longueur de l’organite, qui agirait directement sur l’un ou sur les deux paramètres. Ainsi, le niveau de phosphorylation de la kinase CALK (Chlamydomonas aurora-like protein kinase) chez Chlamydomonas augmente au fur et à mesure que le flagelle s’allonge [34]. Bien qu’aucun lien avec l’IFT n’ait encore été démontré, on pourrait imaginer que cette kinase limite, par exemple, le chargement de la tubuline et des autres précurseurs axonémaux sur les trains IFT.
Les ciliopathies : quand la taille compte
La construction et le maintien de la longueur du cil et du flagelle sont indispensables à la réalisation de leur fonction motrice ou sensorielle. Si la longueur de l’organite n’est pas correcte, la fonction du flagelle n’est que partiellement assurée, et de nombreux organes peuvent être affectés. Les maladies génétiques associées à des défauts ciliaires sont appelées ciliopathies [42] (→). Un point commun à beaucoup de ces pathologies est la modification de la longueur du cil primaire dans les cellules de certains tissus : celui-ci est souvent trop court, mais parfois aussi trop long. Cela pourrait avoir des conséquences directes sur la concentration et la distribution de certains éléments sensoriels, ou sur les propriétés mécaniques du cil, par exemple lorsqu’il doit se plier pour détecter un flux.
(→) Voir la Synthèse de R. Bachmann-Gagescu, page 1011 de ce numéro
Les ciliopathies sont caractérisées par des défauts du développement embryonnaire, ou par des symptômes se manifestant au cours de l’enfance ou de l’adolescence. Les phénotypes cliniques sont variés et affectent nombre d’organes, ce qu’explique la présence quasi ubiquitaire des cils : atteinte rénale avec formation de kystes, encombrement bronchique, infertilité masculine, obésité, rétinopathie pigmentaire, surdité, retard mental, etc. [35, 42] (→).
(→) Voir la Synthèse de R. Bachmann-Gagescu, page 1011 de ce numéro
Le lien entre ciliopathies et cils primaires n’a été établi qu’au début des années 2000, lorsque Greg Pazour et al. ont découvert que le gène Tg737, muté chez des souris modèles de la polykystose rénale (PKD), codait pour la protéine IFT88, essentielle au transport intraflagellaire [22]. Les souris porteuses de la mutation possèdent des cils qui sont trop courts et ne fonctionnent pas normalement. Plus tard, il a été établi que les polycystines, les protéines le plus fréquemment mutées dans cette pathologie, étaient localisées dans ces cils [36]. Des études de génomique comparative ont établi le lien entre plusieurs maladies génétiques et le cil au cours de la dernière décennie [35]. Par exemple, la majorité des gènes impliqués dans le syndrome de Bardet-Biedl (BBS) [43] (→) ne sont conservés que chez les espèces ciliées.
(→) Voir la Synthèse de K. Chennen et al., page 1034 de ce numéro
Les gènes codant pour les protéines du transport intraflagellaire peuvent eux-mêmes être mutés, entraînant la production d’une protéine tronquée ou modifiée. Cette observation suggère que l’absence d’IFT serait létale, comme cela a été démontré chez la souris [37]. Les mutations touchent essentiellement les gènes codant pour les protéines du complexe IFT-A et les moteurs de la chaîne lourde des dynéines. Elles conduisent à des ciliopathies dites « squelettiques », telles que les syndromes de Jeune (caractérisé par un thorax étroit, des membres courts), de Mainzer-Saldino (caractérisé par l’association d’une maladie rénale, d’une dystrophie pigmentaire de la rétine, et d’une dysplasie osseuse) ou de Sensenbrenner (caractérisé par des anomalies squelettiques et ectodermiques associées à une dysmorphie, une néphronophtise, une fibrose hépatique et des anomalies oculaires) [42]. Les cellules des patients construisent toujours des cils, mais la longueur ou la composition de ceux-ci sont modifiées [38, 39].
Conclusion
Ce sont des études sur des organismes modèles (Chlamydomonas, Trypanosoma, Caenorhabditis elegans) qui ont permis d’identifier le transport intraflagellaire et son rôle dans la formation des cils et le contrôle de leur longueur. Plusieurs questions restent encore en suspens. Qu’est-ce qui régule le nombre de trains dans le flagelle ? Qu’est-ce qui contrôle le chargement des trains ? Les mutations observées chez l’homme peuvent maintenant être reproduites chez des organismes modèles, permettant l’analyse de leurs conséquences sur le transport intraflagellaire et le chargements des trains IFT. On peut espérer que ces études permettront de comprendre l’impact des mutations et de discriminer ces dernières des polymorphismes. Ainsi, la combinaison de la recherche fondamentale et de la clinique devrait permettre de réaliser des progrès prometteurs pour le diagnostic des ciliopathies, comme pour la compréhension du mode de formation des cils et flagelles.
Liens d’intérêt
Les auteurs déclarent n’avoir aucun lien d’intérêt concernant les données publiées dans cet article.
Remerciements
Les auteurs remercient Eloïse Bertiaux, Serge Bonnefoy, Sylvie Perrot, Brice Rotureau, Julien Santi-Rocca et Christelle Travaillé pour leurs commentaires sur le manuscrit. Les projets de recherche dans le laboratoire des auteurs sont financés par l’Institut Pasteur, le CNRS et un contrat ANR 11-BSV8-016. Cécile Fort est financée par une bourse du ministère de l’Éducation nationale.
Microscope permettant de visualiser un objet avec un relief donné en exploitant les interférences de deux faisceaux d’une onde lumineuse traversant un échantillon.
On ignore à l’heure actuelle si la modification porte sur la longueur ou la largeur des trains, les observations se fondant sur la détection d’une quantité de fluorescence.
On désigne par précurseurs les composants de l’axonème avant assemblage.
Références
- Bloodgood RA. From central to rudimentary to primary: the history of an underappreciated organelle whose time has come. The primary cilium. Methods Cell Biol 2009 ; 94 : 3–52. [PubMed] [Google Scholar]
- Kohl L, Bastin P. The flagellum of trypanosomes. Int Rev Cytol 2005 ; 224 : 227–285. [CrossRef] [PubMed] [Google Scholar]
- Broadhead R, Dawe HR, Farr H, et al. Flagellar motility is required for the viability of the bloodstream trypanosome. Nature 2006 ; 440 : 224–227. [CrossRef] [PubMed] [Google Scholar]
- Pazour GJ, Agrin N, Leszyk J, Witman GB. Proteomic analysis of a eukaryotic cilium. J Cell Biol 2005 ; 170 : 103–113. [CrossRef] [PubMed] [Google Scholar]
- Ostrowski LE, Blackburn K, Radde KM, et al. A proteomic analysis of human cilia: identification of novel components. Mol Cell Proteomics 2002 ; 1 : 451–465. [CrossRef] [PubMed] [Google Scholar]
- Beisson J, Wright M. Basal body/centriole assembly and continuity. Curr Opin Cell Biol 2003 ; 15 : 96–104. [CrossRef] [PubMed] [Google Scholar]
- Deane JA, Cole DG, Seeley ES, et al. Localization of intraflagellar transport protein IFT52 identifies basal body transitional fibers as the docking site for IFT particles. Curr Biol 2001 ; 11 : 1586–1590. [CrossRef] [PubMed] [Google Scholar]
- Gilula NB, Satir P. The ciliary necklace. A ciliary membrane specialization. J Cell Biol 1972 ; 53 : 494–509. [CrossRef] [PubMed] [Google Scholar]
- Hu Q, Milenkovic L, Jin H, et al. A septin diffusion barrier at the base of the primary cilium maintains ciliary membrane protein distribution. Science 2010 ; 329 : 436–439. [CrossRef] [PubMed] [Google Scholar]
- Rosenbaum JL, Moulder JE, Ringo DL. Flagellar elongation and shortening in Chlamydomonas. The use of cycloheximide and colchicine to study the synthesis and assembly of flagellar proteins. J Cell Biol 1969 ; 41 : 600–619. [CrossRef] [PubMed] [Google Scholar]
- Kozminski KG, Johnson KA, Forscher P, Rosenbaum JL. A motility in the eukaryotic flagellum unrelated to flagellar beating. Proc Natl Acad Sci USA 1993 ; 90 : 5519–5523. [CrossRef] [Google Scholar]
- Pigino G, Geimer S, Lanzavecchia S, et al. Electron-tomographic analysis of intraflagellar transport particle trains in situ. J Cell Biol 2009 ; 187 : 135–148. [CrossRef] [PubMed] [Google Scholar]
- Kozminski KG, Beech PL, Rosenbaum JL. The Chlamydomonas kinesin-like protein FLA10 is involved in motility associated with the flagellar membrane. J Cell Biol 1995 ; 131 : 1517–1527. [CrossRef] [PubMed] [Google Scholar]
- Pazour GJ, Wilkerson CG, Witman GB. A dynein light chain is essential for the retrograde particle movement of intraflagellar transport (IFT). J Cell Biol 1998 ; 141 : 979–992. [CrossRef] [PubMed] [Google Scholar]
- Signor D, Wedaman KP, Orozco JT, et al. Role of a class DHC1b dynein in retrograde transport of IFT motors and IFT raft particles along cilia, but not dendrites, in chemosensory neurons of living Caenorhabditis elegans. J Cell Biol 1999 ; 147 : 519–530. [CrossRef] [PubMed] [Google Scholar]
- Cole DG, Diener DR, Himelblau AL, et al. Chlamydomonas kinesin-II-dependent intraflagellar transport (IFT): IFT particles contain proteins required for ciliary assembly in Caenorhabditis elegans sensory neurons. J Cell Biol 1998 ; 141 : 993–1008. [CrossRef] [PubMed] [Google Scholar]
- Piperno G, Mead K. Transport of a novel complex in the cytoplasmic matrix of Chlamydomonas flagella. Proc Natl Acad Sci USA 1997 ; 94 : 4457–4462. [CrossRef] [Google Scholar]
- Taschner M, Bhogaraju S, Lorentzen E. Architecture and function of IFT complex proteins in ciliogenesis. Differentiation 2012 ; 83 : S12–S22. [CrossRef] [PubMed] [Google Scholar]
- Orozco JT, Wedaman KP, Signor D, et al. Movement of motor, cargo along cilia. Nature 1999 ; 398 : 674. [CrossRef] [PubMed] [Google Scholar]
- Brown JM, Marsala C, Kosoy R, Gaertig J. Kinesin-II is preferentially targeted to assembling cilia and is required for ciliogenesis and normal cytokinesis in Tetrahymena. Mol Biol Cell 1999 ; 10 : 3081–3096. [CrossRef] [PubMed] [Google Scholar]
- Absalon S, Blisnick T, Kohl L, et al. Intraflagellar transport and functional analysis of genes required for flagellum formation in trypanosomes. Mol Biol Cell 2008 ; 19 : 929–944. [CrossRef] [PubMed] [Google Scholar]
- Pazour GJ, Dickert BL, Vucica Y, et al. Chlamydomonas IFT88 and its mouse homologue, polycystic kidney disease gene Tg737, are required for assembly of cilia and flagella. J Cell Biol 2000 ; 151 : 709–718. [CrossRef] [PubMed] [Google Scholar]
- Follit JA, Tuft RA, Fogarty KE, Pazour GJ. The intraflagellar transport protein IFT20 is associated with the Golgi complex and is required for cilia assembly. Mol Biol Cell 2006 ; 17 : 3781–3792. [CrossRef] [PubMed] [Google Scholar]
- Buisson J, Chenouard N, Lagache T, et al. Intraflagellar transport proteins cycle between the flagellum and its base. J Cell Sci 2013 ; 126 : 327–338. [CrossRef] [PubMed] [Google Scholar]
- Nonaka S, Tanaka Y, Okada Y, et al. Randomization of left-right asymmetry due to loss of nodal cilia generating leftward flow of extraembryonic fluid in mice lacking KIF3B motor protein. Cell 1998 ; 95 : 829–837. [CrossRef] [PubMed] [Google Scholar]
- Kohl L, Robinson D, Bastin P. Novel roles for the flagellum in cell morphogenesis and cytokinesis of trypanosomes. EMBO J 2003 ; 22 : 5336–5346. [CrossRef] [PubMed] [Google Scholar]
- Bhogaraju S, Cajanek L, Fort C, et al. Molecular basis of tubulin transport within the cilium by IFT74 and IFT81. Science 2013 ; 341 : 1009–1012. [CrossRef] [PubMed] [Google Scholar]
- Marshall WF, Rosenbaum JL. Intraflagellar transport balances continuous turnover of outer doublet microtubules: implications for flagellar length control. J Cell Biol 2001 ; 155 : 405–414. [CrossRef] [PubMed] [Google Scholar]
- Dentler W. Intraflagellar transport (IFT) during assembly and disassembly of Chlamydomonas flagella. J Cell Biol 2005 ; 170 : 649–659. [CrossRef] [PubMed] [Google Scholar]
- Engel BD, Ludington WB, Marshall WF. Intraflagellar transport particle size scales inversely with flagellar length: revisiting the balance-point length control model. J Cell Biol 2009 ; 187 : 81–89. [CrossRef] [PubMed] [Google Scholar]
- Wren KN, Craft JM, Tritschler D, et al. A differential cargo-loading model of ciliary length regulation by IFT. Curr Biol 2013 ; 23 : 2463–2471. [CrossRef] [PubMed] [Google Scholar]
- Besschetnova TY, Kolpakova-Hart E, Guan Y, et al. Identification of signaling pathways regulating primary cilium length and flow-mediated adaptation. Curr Biol 2010 ; 20 : 182–187. [CrossRef] [PubMed] [Google Scholar]
- Pan J, Snell WJ. Chlamydomonas shortens its flagella by activating axonemal disassembly, stimulating IFT particle trafficking, and blocking anterograde cargo loading. Dev Cell 2005 ; 9 : 431–438. [CrossRef] [PubMed] [Google Scholar]
- Cao M, Meng D, Wang L, et al. Activation loop phosphorylation of a protein kinase is a molecular marker of organelle size that dynamically reports flagellar length. Proc Natl Acad Sci USA 2013 ; 110 : 12337–12342. [CrossRef] [Google Scholar]
- Hildebrandt F, Benzing T, Katsanis N. Ciliopathies. N Engl J Med 2011 ; 364 : 1533–1543. [CrossRef] [PubMed] [Google Scholar]
- Pazour GJ, San Agustin JT, Follit JA, et al. Polycystin-2 localizes to kidney cilia and the ciliary level is elevated in orpk mice with polycystic kidney disease. Curr Biol 2002 ; 12 : R378–R380. [CrossRef] [PubMed] [Google Scholar]
- Huangfu D, Liu A, Rakeman AS, et al. Hedgehog signalling in the mouse requires intraflagellar transport proteins. Nature 2003 ; 426 : 83–87. [CrossRef] [PubMed] [Google Scholar]
- Walczak-Sztulpa J, Eggenschwiler J, Osborn D, et al. Cranioectodermal dysplasia, Sensenbrenner syndrome, is a ciliopathy caused by mutations in the IFT122 gene. Am J Hum Genet 2010 ; 86 : 949–956. [CrossRef] [PubMed] [Google Scholar]
- Perrault I, Saunier S, Hanein S, et al. Mainzer-Saldino syndrome is a ciliopathy caused by IFT140 mutations. Am J Hum Genet 2012 ; 90 : 864–870. [CrossRef] [PubMed] [Google Scholar]
- Vincensini L, Blisnick T, Bastin P. 1001 model organisms to study cilia and flagella. Biol Cell 2011 ; 103 : 109–130. [CrossRef] [PubMed] [Google Scholar]
- Buisson J, Bastin P. Flagellum structure and function in trypanosomes. Microbiol Monogr 2010 ; 17 : 63–86. [CrossRef] [Google Scholar]
- Bachmann-Gagescu R. Complexité génétique des ciliopathies et identification de nouveaux gènes. Med Sci (Paris) 2014 ; 30 : 1011–1023. [CrossRef] [EDP Sciences] [PubMed] [Google Scholar]
- Chennen K, Scerbo MJ, Dollfus H, et al. Syndrome de Bardet-Biedl : cils et obésité. De la génétique à l’approche intégrative. Med Sci (Paris) 2014 ; 30 : 1034–1039. [CrossRef] [EDP Sciences] [PubMed] [Google Scholar]
Liste des figures
|
Figure 1. Diversité du nombre, de la longueur et de la disposition des cils. A. Épithélium tapissant les ventricules du cerveau de la souris. B. Épithélium du tube collecteur du rein de souris visualisé après cryofracture mécanique. C. Cil primaire à la surface de cellules IMCD (inner medullary collecting duct) et ( D ) de cellules MDCK (Mardin Darby canine kidney). Toutes les images ont été prises au microscope électronique à balayage et ont été modifiées à partir de Vincensini et al. [40]. Barres d’échelle : 1 µm. Les cils sont indiqués par des flèches. |
| Dans le texte | |
|
Figure 2. Ultrastructure du flagelle de Trypanosoma brucei. A. Coupe longitudinale de la base du flagelle. Le corps basal immature n’est pas visible sur cette section. Les mentions B-E dans le panneau A indiquent les sections transversales à travers le flagelle dont les champs sont grossis dans la colonne de droite de la figure. B. Corps basal. C. Le début de la zone de transition avec les fibres de transition dont l’extrémité est indiquée par des flèches. D. La portion centrale de la zone de transition. Les têtes de flèches indiquent la collerette. E. Axonème dans la lumière de la poche flagellaire. Du matériel dense aux électrons est fréquemment observé entre les doublets périphériques et la membrane. F. Représentation schématique de la base du flagelle indiquant le corps basal (9 triplets de microtubules), la zone de transition (9 doublets de microtubules) et l’axonème (9 doublets de microtubules représentés en bleu foncé et une paire de singulets représentés en bleu clair). FPC : collier de la poche flagellaire ; K : kinétoplaste ; G : appareil de Golgi. Barres d’échelle : 100 nm. Les photographies sont extraites de [41]. |
| Dans le texte | |
|
Figure 3. Modèle du transport intraflagellaire. (1) Les particules IFT, les moteurs et les précurseurs axonémaux sont recrutés à la base du flagelle. (2) L’assemblage des particules IFT et le chargement des précurseurs axonémaux auraient lieu au niveau des fibres de transition (représentées en noir). (3) La kinésine II et le complexe IFT-B sont indispensables au transport antérograde en direction de la pointe du flagelle. La dynéine est transportée sous une forme inactive. (4) Les particules IFT sont remaniées à l’extrémité distale du flagelle. (5) Les particules retournent à la base du flagelle par l’intermédiaire de la dynéine, responsable du transport rétrograde, pour y être recyclées [24]. La kinésine est alors inactivée et revient vers la base soit sous forme de cargo [15], soit par diffusion [30]. KAP : kinesin accessory protein ; DIC5 : chaîne intermédiaire de dynéine ; DLIC1 : chaîne légère intermédiaire de dynéine ; DHC2.1 : chaîne lourde de dynéine 2. |
| Dans le texte | |
|
Figure 4. Modèle de l’équilibre du nombre de particules IFT. Les points verts représentent les protéines axonémales. A. Au stade précoce de formation du cil, l’apport de tubuline est supérieur à la perte et le flagelle s’allonge. B. Lorsque le cil grandit, le taux de croissance ralentit à cause de l’augmentation de la distance à parcourir jusqu’à atteindre un équilibre entre assemblage et désassemblage de la tubuline. |
| Dans le texte | |
|
Figure 5. Modèle du contrôle de la longueur du cil par la quantité de protéines IFT disponibles dans l’organite. A. Une quantité fixe d’IFT est injectée dans le cil au stade précoce de sa formation. Les trains sont alors longs et peuvent transporter une grande quantité de précurseurs comme la tubuline dont l’apport est supérieur à la perte, permettant ainsi l’élongation du cil. B. Lorsque le cil s’allonge, les trains deviennent plus petits et apportent de moins en moins de précurseurs au bout distal, jusqu’à atteindre un équilibre dynamique. |
| Dans le texte | |
|
Figure 6. Modèle du contrôle de la longueur du flagelle en fonction du chargement des précurseurs axonémaux sur les particules IFT. A. Au stade précoce de construction, le chargement de précurseurs axonémaux est important et permet au cil de s’allonger rapidement. B. Le taux de chargement se réduit lorsque le flagelle s’allonge, jusqu’à atteindre l’équilibre entre assemblage et désassemblage. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.