")
")
| Issue |
Med Sci (Paris)
Volume 29, Number 4, Avril 2013
|
|
|---|---|---|
| Page(s) | 355 - 358 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/2013294007 | |
| Published online | 26 avril 2013 | |
Une avancée technique pour comprendre la dynamique de l’horloge de segmentation
A technical breakthrough for understanding segmentation clock dynamics and synchrony
1
Laboratoire de biologie moléculaire de la cellule (LBMC), UMR5239, CNRS/UCBL/ENS de Lyon, 46, allée d’Italie, 69007 Lyon, France
2
McGill university, Montreal, Québec, Canada
3
University of California at Berkeley, Berkeley, États-Unis
4
The Ohio state university, Columbus, États-Unis
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Chez les vertébrés, certains éléments du corps, tels que les vertèbres et les muscles axiaux, sont répétés le long de l’axe antéropostérieur. Cette métamérie prend naissance pendant le développement embryonnaire lors de la segmentation du mésoderme présomitique. Ce dernier s’étend longitudinalement de part et d’autre du plan médian de l’embryon (Figure 1A). Au fur et à mesure que le mésoderme présomitique s’allonge grâce à l’arrivée de nouvelles cellules à son extrémité postérieure, les cellules les plus antérieures se séparent périodiquement du mésoderme présomitique et forment des blocs de cellules épithéliales, appelés somites. La somitogenèse constitue un magnifique exemple de structuration d’un tissu au cours du développement, qui frappe par son extraordinaire périodicité, à la fois spatiale et temporelle. Comment cette périodicité est-elle contrôlée ?
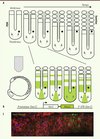 |
Figure 1. Stratégie de visualisation en temps réel de l’activité de l’horloge de segmentation. A. Modèle de somitogenèse. Le mésoderme présomitique (PSM) et les somites sont schématisés en vue dorsale. Les flèches grises représentent l’arrivée continuelle de nouvelles cellules dans le mésoderme présomitique postérieur, et le point noir, une de ces cellules. Au fur et à mesure de la segmentation, cette cellule devient de plus en plus antérieure au sein du mésoderme présomitique, alors que sa position absolue par rapport à l’axe antéropostérieur de l’embryon varie peu. La transcription d’un gène cyclique typique au cours d’un cycle de segmentation est figurée en vert. B. Rapporteur de l’horloge, basé sur les séquences codante et régulatrices du gène cyclique her1. C. Image en microscopie confocale du mésoderme présomitique (vue latérale) dans un embryon exprimant le rapporteur de l’horloge Her1-Venus (vert), ainsi que des marqueurs membranaire (rouge) et nucléaire (bleu). Le signal nucléaire Her1-Venus est détecté avec un niveau de résolution cellulaire. |
Analyse par un système rapporteur des oscillations de l’horloge au cours de la somitogenèse
La somitogenèse est régie par un oscillateur moléculaire, l’horloge de segmentation, dont l’existence est révélée par la transcription périodique de gènes « cycliques » dans le mésoderme présomitique [1]. Des vagues d’expression de ces gènes cycliques sont générées dans le mésoderme présomitique postérieur avec une période identique à la celle de la somitogenèse et se propagent antérieurement (Figure 1A). L’horloge de segmentation reposerait sur un réseau complexe d’interactions génétiques et biochimiques, dont la nature n’est pas encore bien comprise, et qui implique une coordination de la multitude d’oscillateurs individuels que sont les cellules du mésoderme présomitique.
La dynamique de l’horloge a été abordée par diverses approches [2] : analyse du profil d’expression des gènes cycliques dans des cohortes d’embryons fixés, comparaison de ce profil entre des moitiés d’embryon dont le développement est à un stade d’avancement différent, marquage à haute résolution permettant de distinguer les transcrits naissants (nucléaires) des transcrits tardifs (cytoplasmiques) et, plus récemment, imagerie directe en temps réel de rapporteurs permettant de visualiser l’expression de gènes cycliques chez la souris. Pour comprendre les mécanismes qui génèrent et synchronisent les oscillations, il est essentiel de visualiser l’activité de l’horloge au niveau cellulaire. Visualiser un processus aussi dynamique grâce à un système rapporteur est délicat car : (1) le rapporteur doit être produit et dégradé dans le court temps d’un cycle de l’horloge (2 h chez la souris, 30 min chez le poisson zèbre), (2) le rapporteur doit rester détectable malgré sa grande instabilité, et (3) l’activité de l’horloge endogène ne doit pas être affectée par le rapporteur [2]. Les premiers rapporteurs de l’horloge développés chez la souris ont permis de visualiser les vagues d’expression de gènes cycliques au niveau tissulaire [3–5]. Afin d’atteindre un niveau de résolution cellulaire, nous nous sommes tournés vers le modèle du poisson zèbre, dont la transparence et le développement externe sont des atouts pour l’imagerie à haute résolution. Dans un travail publié récemment dans Developmental Cell, nous avons développé un système rapporteur très dynamique de l’horloge, ainsi que des outils essentiels de mesure des oscillations, et nous avons analysé les mécanismes de synchronisation des cellules du mésoderme présomitique [6].
Utilisation du rapporteur Her1-Venus et mesure des oscillations de l’horloge à l’échelle cellulaire
Notre système rapporteur est basé sur les séquences régulatrices (promoteur et 3’ UTR) du gène cyclique her1 (hairy/enhancer of split-related gene) [7]. Lorsque la séquence codant pour la protéine GFP (green fluorescent protein) est placée sous le contrôle du promoteur her1, elle est transcrite de manière cyclique, mais produit un signal uniforme en raison de sa grande stabilité [7]. Nous avons utilisé Venus, une protéine dérivée de la GFP dont la maturation est rapide, et nous avons testé plusieurs stratégies pour en diminuer la demi-vie. En fusionnant Venus avec une séquence PEST (séquence riche en proline [P], acide glutamique [E], sérine [S], et thréonine [T]) ou un motif bi-ubiquitine, deux signaux de dégradation protéique utilisés l’un et l’autre avec succès pour générer des rapporteurs de l’horloge chez la souris [3, 4], nous avons obtenu des rapporteurs soit trop stables (motif PEST), soit au contraire trop instables pour être détectés en temps réel (motif bi-ubiquitine). En revanche, nous avons observé des oscillations en fusionnant Venus avec la séquence codante de la protéine Her1, elle-même instable (Figure 1B). Nous avons établi une lignée transgénique de poissons zèbre exprimant Her1-Venus et montré que l’impact du rapporteur sur l’horloge endogène est minime dans des embryons hétérozygotes.
Pour mesurer l’évolution du signal Her1-Venus dans des cellules du mésoderme présomitique, nous avons filmé en microscopie confocale des embryons exprimant non seulement le rapporteur, mais aussi des marqueurs membranaire et nucléaire des cellules (Figure 1C). Nous avons développé un programme pour délimiter les cellules dans les trois dimensions, les suivre au cours du temps et quantifier à chaque instant le signal Her1-Venus nucléaire. Les paramètres des oscillations ont été extraits des courbes de mesure brutes à l’aide d’un algorithme validé empiriquement. En particulier, nous avons mesuré dans quelle phase du cycle d’oscillation se trouve chaque cellule à un instant donné.
Importance de la voie Notch pour la synchronisation des oscillations
La coloration virtuelle des noyaux en fonction de la phase des cellules permet de visualiser directement un ralentissement des oscillations le long de l’axe antéropostérieur, caractéristique des vagues d’expression des gènes cycliques (Figure 2A). Le caractère progressif de cette transition de phase indique que les cellules voisines oscillent de manière synchronisée, ce que nous avons pu quantifier en calculant la différence de phase sur un grand nombre de couples de cellules voisines. Comme la segmentation est très perturbée dans les mutants de la voie de signalisation Notch, et que les gènes cycliques présentent un profil d’expression erratique dans le mésoderme présomitique d’embryons fixés, il a été suggéré en 2000 que les cellules oscillent de manière désynchronisée en l’absence de signalisation Notch [8]. En mesurant l’activité de Her1-Venus dans divers mutants de la voie Notch, nous avons pu confirmer cette interprétation au moyen d’une démonstration directe (Figure 2B).
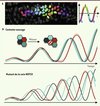 |
Figure 2. Impact de la voie de signalisation Notch et de la mitose sur la synchronisation de cellules voisines. A. Image de cellules du mésoderme présomitique (en vue latérale, antérieur à droite) dont les noyaux sont colorés en fonction de leur phase d’oscillation. Dans cet embryon exprimant Her1-Venus en contexte sauvage, les cellules présentent une transition de phase progressive le long de l’axe antéropostérieur, indiquant que leurs oscillations sont finement coordonnées. B. Bilan schématisé de nos résultats. Un groupe de cellules voisines oscillent de manière synchrone dans des embryons exprimant Her1-Venus en contexte sauvage. Elles oscillent encore, mais de manière désynchronisée, dans des mutants de la voie Notch. Dans des contextes sauvage ou mutant de la voie Notch, la division cellulaire perturbe les oscillations et génère deux cellules filles parfaitement synchronisées (qui se resynchronisent ensuite progressivement avec leurs voisines dans les embryons sauvages). |
Impact de la mitose sur les oscillations de l’horloge
Le maintien de la synchronisation est essentiel pour lutter contre des perturbations d’oscillations qui peuvent survenir notamment lors de la division cellulaire [9]. Nous avons observé un arrêt temporaire des oscillations dans la plupart des cas de mitose. À la reprise des oscillations, les cellules sœurs issues d’une même mitose sont généralement déphasées par rapport à leurs voisines mais, de manière surprenante, elles restent extrêmement synchronisées entre elles, que ce soit dans des contextes sauvage ou mutant de la voie Notch (Figure 2B). Ceci suggère que chaque cellule hériterait d’un ensemble de facteurs suffisants pour déterminer le démarrage de plusieurs oscillations, et souligne la robustesse d’un mécanisme intracellulaire et indépendant de Notch qui génère et/ou entretient les oscillations. Enfin, nous avons observé que les mitoses surviennent préférentiellement lorsque le groupe de cellules environnantes se trouve dans une certaine phase du cycle d’oscillation. Ainsi, l’horloge serait possiblement liée au cycle cellulaire via des mécanismes qui restent à élucider.
Conclusions et perspectives
Notre travail ouvre la porte à une nouvelle approche pour étudier les mécanismes de l’horloge. Bien que Her1-Venus présente certaines limitations, telles qu’un signal dans le mésoderme présomitique postérieur trop faible pour analyser les mécanismes d’initiation et de synchronisation des cellules dans cette région, il est possible d’obtenir un système rapporteur à la fois hautement instable et détectable chez le poisson zèbre. Les progrès réalisés dans la synthèse de protéines fluorescentes dont la maturation est rapide, ainsi que dans la sensibilité des techniques d’imagerie, permettront de continuer à développer de nouveaux rapporteurs. En outre, les outils de détection et de suivi des cellules, ainsi que l’analyse des oscillations permettront de développer le même type d’approche chez d’autres vertébrés.
Liens d’intérêt
Les auteurs déclarent n’avoir aucun lien d’intérêt concernant les données publiées dans cet article.
Références
- Pourquié O. Vertebrate segmentation: from cyclic gene networks to scoliosis. Cell 2011 ; 145 : 650–663. [CrossRef] [PubMed] [Google Scholar]
- Soroldoni D, Oates AC. Live transgenic reporters of the vertebrate embryo’s segmentation clock. Curr Opin Genet Dev 2011 ; 21 : 600–605. [CrossRef] [PubMed] [Google Scholar]
- Masamizu Y, Ohtsuka T, Takashima, Y, et al. Real-time imaging of the somite segmentation clock: revelation of unstable oscillators in the individual presomitic mesoderm cells. Proc Natl Acad Sci USA 2006 ; 103 : 1313–1318. [CrossRef] [Google Scholar]
- Aulehla A, Wiegraebe W, Baubet V, et al. A beta-catenin gradient links the clock and wavefront systems in mouse embryo segmentation. Nat Cell Biol 2008 ; 10 : 186–193. [Google Scholar]
- Takashima Y, Ohtsuka T, Gonzalez A, et al. Intronic delay is essential for oscillatory expression in the segmentation clock. Proc Natl Acad Sci USA 2011 ; 108 : 3300–3305. [CrossRef] [Google Scholar]
- Delaune EA, François P, Shih NP, Amacher SL. Single-cell-resolution imaging of the impact of Notch signaling and mitosis on segmentation clock dynamics. Dev Cell 2012 ; 23 : 995–1005. [CrossRef] [PubMed] [Google Scholar]
- Gajewski M, Sieger D, Alt B, et al. Anterior and posterior waves of cyclic her1 gene expression are differentially regulated in the presomitic mesoderm of zebrafish. Development 2003 ; 130 : 4269–4278. [CrossRef] [PubMed] [Google Scholar]
- Jiang YJ, Aerne BL, Smithers L, et al. Notch signalling and the synchronization of the somite segmentation clock. Nature 2000 ; 408 : 475–479. [CrossRef] [PubMed] [Google Scholar]
- Horikawa K, Ishimatsu K, Yoshimoto E, et al.Noise-resistant and synchronized oscillation of the segmentation clock. Nature 2006 ; 441 : 719–723. [CrossRef] [PubMed] [Google Scholar]
© 2013 médecine/sciences – Inserm / SRMS
Liste des figures
|
Figure 1. Stratégie de visualisation en temps réel de l’activité de l’horloge de segmentation. A. Modèle de somitogenèse. Le mésoderme présomitique (PSM) et les somites sont schématisés en vue dorsale. Les flèches grises représentent l’arrivée continuelle de nouvelles cellules dans le mésoderme présomitique postérieur, et le point noir, une de ces cellules. Au fur et à mesure de la segmentation, cette cellule devient de plus en plus antérieure au sein du mésoderme présomitique, alors que sa position absolue par rapport à l’axe antéropostérieur de l’embryon varie peu. La transcription d’un gène cyclique typique au cours d’un cycle de segmentation est figurée en vert. B. Rapporteur de l’horloge, basé sur les séquences codante et régulatrices du gène cyclique her1. C. Image en microscopie confocale du mésoderme présomitique (vue latérale) dans un embryon exprimant le rapporteur de l’horloge Her1-Venus (vert), ainsi que des marqueurs membranaire (rouge) et nucléaire (bleu). Le signal nucléaire Her1-Venus est détecté avec un niveau de résolution cellulaire. |
| Dans le texte | |
|
Figure 2. Impact de la voie de signalisation Notch et de la mitose sur la synchronisation de cellules voisines. A. Image de cellules du mésoderme présomitique (en vue latérale, antérieur à droite) dont les noyaux sont colorés en fonction de leur phase d’oscillation. Dans cet embryon exprimant Her1-Venus en contexte sauvage, les cellules présentent une transition de phase progressive le long de l’axe antéropostérieur, indiquant que leurs oscillations sont finement coordonnées. B. Bilan schématisé de nos résultats. Un groupe de cellules voisines oscillent de manière synchrone dans des embryons exprimant Her1-Venus en contexte sauvage. Elles oscillent encore, mais de manière désynchronisée, dans des mutants de la voie Notch. Dans des contextes sauvage ou mutant de la voie Notch, la division cellulaire perturbe les oscillations et génère deux cellules filles parfaitement synchronisées (qui se resynchronisent ensuite progressivement avec leurs voisines dans les embryons sauvages). |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.