")
")
| Issue |
Med Sci (Paris)
Volume 29, Number 1, Janvier 2013
|
|
|---|---|---|
| Page(s) | 75 - 81 | |
| Section | M/S Revues | |
| DOI | https://doi.org/10.1051/medsci/2013291016 | |
| Published online | 25 janvier 2013 | |
L’intestin moyen de drosophile
Un nouveau modèle d’étude des cellules souches adultes in vivo
The Drosophila midgut as a model to study adult stem cells
Génétique et biologie du développement, Institut Curie, UMR 3215 CNRS, Inserm U934, 26, rue d’Ulm, 75248 Paris Cedex 05, France
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Résumé
Le renouvellement des organes est assuré tout au long de la vie d’un individu par des cellules souches adultes spécifiques à chaque tissu. L’identification de cellules souches adultes actives au sein de l’intestin de drosophile a permis de développer un modèle prometteur d’étude des cellules souches adultes in vivo. Nous détaillons ici les mécanismes qui contrôlent le choix du destin cellulaire dans le lignage intestinal en développant plus précisément le rôle de la voie Notch. Nous décrivons également comment le contrôle de la prolifération des cellules souches intestinales est crucial pour le maintien de l’homéostasie tissulaire au cours du renouvellement naturel du tissu et en réponse au stress environnemental.
Abstract
Constant renewal of cells occurs in most tissues throughout the adult lifetime and is insured by the activity of resident stem cells. Recent work has demonstrated the presence of adult stem cells in the Drosophila intestine and consequently, the Drosophila intestine has become a powerful model to understand adult stem cells in vivo. In this review, we summarize our current understanding of the mechanisms controlling cell fate decisions of the intestinal stem cells with a particular focus on the role of the Notch pathway in this process. We also summarize what is known about proliferation control of the intestinal stem cells, which is crucial to maintain tissue homeostasis during normal and environmentally stressful conditions.
© 2013 médecine/sciences – Inserm / SRMS
Les cellules souches adultes
Les cellules souches ont un rôle essentiel, non seulement au cours du développement, mais également pendant la vie adulte. Elles ont comme caractéristique principale de pouvoir se diviser pour produire de nouvelles cellules souches (autorenouvellement) et des cellules capables de se spécialiser (différenciation). Grâce à cette propriété, les cellules souches adultes renouvellent nos tissus et assurent leur réparation suite à d’éventuelles lésions.
Comprendre comment l’équilibre entre la prolifération et la différenciation des cellules souches adultes est contrôlé est devenu une question cruciale de la recherche biomédicale. En effet, un défaut dans le maintien du nombre de cellules souches pourrait être à l’origine de la dégénérescence tissulaire liée au vieillissement et du développement de certains cancers [1, 2]. De plus, les cellules souches possèdent des caractéristiques communes avec les cellules souches cancéreuses potentiellement impliquées dans le développement tumoral [3]. Enfin, l’utilisation de cellules souches adultes comme source de cellules différenciées ouvre de nouvelles perspectives médicales en termes de régénération ou de remplacement tissulaire [4].
La drosophile : un modèle d’étude des cellules souches in vivo
L’utilisation de la drosophile (Drosophila melanogaster) comme organisme modèle permet d’étudier le comportement des cellules souches in vivo au sein de tissus simples, tout en disposant de nombreux outils génétiques. Des avancées significatives dans la connaissance des mécanismes régulant les cellules souches et conservés chez les mammifères ont ainsi été réalisées chez la drosophile. L’étude des cellules souches germinales a mis en évidence le rôle du microenvironnement dans le contrôle du devenir des cellules souches et a permis de valider le concept de niche [5, 6]. De même, le processus de division des neuroblastes (cellules progénitrices neurales partageant un grand nombre de caractéristiques avec les cellules souches) est un excellent exemple de divisions asymétriques contrôlant le renouvellement et la différenciation des cellules souches [7].
Ces dernières années, différentes populations de cellules souches adultes ont été identifiées au sein des voies digestives chez la drosophile par l’analyse de lignages cellulaires (étude de la descendance issue d’une même cellule). Ces cellules sont capables de se maintenir tout au long de la vie de l’individu et permettent le renouvellement de l’ensemble des cellules différenciées du tissu [8, 9]. L’intestin de drosophile (Figure 1A) est un tissu en renouvellement constant, capable de se régénérer rapidement en réponse au stress environnemental. Il est ainsi devenu un modèle privilégié d’étude du maintien de l’homéostasie tissulaire par des cellules souches adultes.
 |
Figure 1. Organisation structurelle et cellulaire de l’intestin moyen de drosophile. A. Le système digestif est connecté en son extrémité antérieure à la bouche, aux glandes salivaires (non représentées) et au jabot. Il se prolonge jusqu’à l’intestin antérieur composé, entre autres, de l’œsophage. Ce dernier aurait une fonction similaire à celle de l’estomac. L’appareil digestif est également composé d’un intestin moyen et d’un intestin postérieur. L’intestin moyen de drosophile est segmenté en trois parties majeures : une région antérieure (rouge), une région intermédiaire (vert) qui inclut une zone composée de cellules Cu2+ au pH très acide (pH < 3), et une région postérieure (bleu). B. L’épithélium intestinal est entouré de cellules musculaires viscérales (MV), et est composé de cellules différenciées : les cellules entéroendocrines (ee, cellules sécrétrices) et les entérocytes (EC, cellules absorbantes), et de cellules progénitrices : les cellules souches intestinales (CSI) et les entéroblastes (EB, cellules post-mitotiques). C. Le lignage intestinal cellulaire est simple. Les cellules souches intestinales sont les seules cellules capables de se diviser et la majorité d’entre elles génèrent, après division, une nouvelle cellule souche et un entéroblaste. Cet entéroblaste a la capacité de poursuivre sa différenciation pour devenir soit une cellule entéroendocrine (ee), soit un entérocyte (EC). |
Dans cette revue, nous présentons comment le destin cellulaire est déterminé au sein du lignage intestinal et, en particulier, comment la voie Notch joue un rôle majeur dans ce processus. Dans un deuxième temps, nous nous intéresserons à la régulation de la prolifération cellulaire en condition d’homéostasie tissulaire, mais également au cours de la réparation tissulaire générée par un stress environnemental.
Les cellules souches adultes de l’intestin de drosophile
Chez la drosophile, le système digestif peut être séparé en trois parties : l’intestin antérieur connecté à la bouche, aux glandes salivaires et au jabot, l’intestin moyen représentant la majeure partie du tube digestif et l’intestin postérieur comparable au côlon chez les mammifères (Figure 1A). Différentes populations de cellules souches ont été identifiées dans chaque portion du système digestif de drosophile. Cependant, la majorité des travaux de recherche se sont concentrés sur les cellules souches de l’intestin moyen que nous appellerons par la suite « intestin » pour plus de simplicité.
L’intestin de drosophile est constitué d’un mono-épithélium pseudostratifié entouré de cellules musculaires viscérales (Figure 1B). Les différentes fonctions physiologiques du tissu sont assurées par deux types de cellules différenciées : les cellules sécrétrices, appelées cellules entéroendocrines, et les cellules absorbantes, appelées entérocytes. Il existe également deux catégories de cellules progénitrices : les cellules souches intestinales (CSI) et des cellules postmitotiques engagées dans la voie de différenciation appelées entéroblastes. Comme chez les mammifères, l’homéostasie de l’épithélium intestinal repose sur la capacité des CSI multipotentes à renouveler l’ensemble des cellules différenciées du tissu tout au long de la vie de l’individu. Le lignage cellulaire est toutefois plus simple chez la drosophile (Figure 1C) [10–12]. Les CSI sont les seules cellules capables de se diviser au sein du tissu, donnant naissance, dans la majorité des cas, une nouvelle CSI et un entéroblaste. Ce dernier poursuit sa différenciation pour former soit une cellule entéroendocrine, soit un entérocyte (Figure 1C) [8, 9].
La voie Notch est essentielle à la spécification du destin cellulaire
À l’instar de la situation chez les mammifères, la voie de signalisation Notch [13] est essentielle à la spécification du destin cellulaire au sein du lignage intestinal chez la drosophile (Figure 2). Immédiatement après la division de la CSI, le récepteur Notch et son ligand Delta sont présents dans les deux cellules filles. Cependant, la voie Notch ne sera activée que dans une seule des deux cellules qui deviendra alors un entéroblaste. Cette activation est dépendante du ligand Delta présent dans l’autre cellule fille (la nouvelle CSI). On ne sait toutefois pas si la restriction de l’expression de la protéine Delta à la nouvelle cellule souche se fait avant, pendant ou après l’activation de la voie Notch [9, 14].
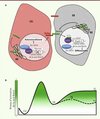 |
Figure 2. Modèle de régulation de la voie Notch dans le lignage intestinal. A. En l’absence d’activation de la voie Notch, Hairless permettrait le maintien des cellules souches (CSI) en réprimant l’expression des gènes cibles de la voie. L’activité du facteur de transcription Daughterless (Da) serait ainsi maintenue entraînant le renouvellement des CSI et le maintien de l’expression de Delta. L’activation de la voie Notch par le ligand Delta dans la cellule voisine entraîne le clivage du récepteur Notch et la translocation de son domaine intracellulaire (NICD) dans le noyau. Le complexe NICD/suppressor of hairless (Su[H]) active la transcription des gènes cibles de la voie Notch, parmi lesquels les gènes codant pour les membres du complexe enhancer of split (E[spl]-C). Le complexe E(spl)-C aurait comme fonction d’inhiber l’activité de Da, permettant ainsi la différenciation de la cellule en entéroblaste (EB). L’activité de la voie Notch est modulée par des modifications post-traductionnelles du récepteur Notch (fucosylation, glycosylation, en jaune). Aucune restriction de ces modifications aux EB n’a été rapportée jusqu’ici. RE : réticulum endoplasmique. B. Un niveau élevé d’activation de la voie Notch dépendant de la fucosylation du récepteur doit être atteint pour entraîner le basculement de la cellule souche dans la voie de différenciation (EB). Dans un deuxième temps, la différenciation terminale des entéroblastes en cellules entérocytes (EC) requiert un niveau plus faible d’activité de la voie Notch, indépendant de la fucosylation du récepteur. La relation temporelle entre ces événements n’est cependant pas caractérisée à l’heure actuelle. Il a également été proposé que différents niveaux d’activation de Notch participent au choix entre les destins des cellules entéroendocrines (ee) et des entérocytes (EC) : un niveau plus élevé serait requis pour former un entérocyte, alors qu’un faible niveau d’activation permettrait la différenciation de l’entéroblaste en cellule entéroendocrine. |
Le rôle de la voie Notch dans le lignage intestinal a pu être évalué en bloquant son activité ou en l’activant de manière ectopique. En absence d’activation de cette voie, des cellules ayant les caractéristiques des CSI et des cellules entéroendocrines s’accumulent massivement au détriment des entérocytes qui sont quasiment absents. À l’inverse, l’expression d’une forme constitutivement active du récepteur Notch dans les CSI entraîne la perte de ces dernières du fait de leur différenciation en entérocytes [8, 9]. La voie Notch a donc un double rôle dans la régulation du lignage intestinal : un premier dans le choix entre l’autorenouvellement et l’engagement dans la différenciation des CSI, et un second dans le choix entre les destins entéroendocrine et entérocyte lors de la différenciation terminale.
L’activation de la voie Notch spécifiquement dans une seule des deux cellules filles est importante afin de maintenir les CSI dans leur état non différencié. Un des mécanismes qui restreint l’activité de cette voie dans les CSI repose sur la répression, par le complexe Hairless/Suppressor of Hairless (Su[H]), de l’expression des gènes cibles de Notch (dont les membres du complexe Enhancer of split [E(spl)-C]) (Figure 2) [15]. Dans les entéroblastes, l’interaction du récepteur Notch avec Delta entraîne le clivage de Notch, suivi de la translocation de son domaine intracellulaire NICD (Notch intracellular domain) dans le noyau. Ce dernier se lie à Su(H) à la place de Hairless, et induit la transcription des gènes cibles de la voie Notch, dont E(spl)-C [16, 17]. Le rôle des protéines E(spl)-C serait d’inhiber l’activité du facteur de transcription Daughterless [18], lui-même nécessaire au renouvellement des CSI, ce qui permettrait l’entrée de la CSI dans la voie de différenciation. Ainsi, une perte de fonction de E(spl)-C induit la persistance de l’expression de Delta et l’accumulation de cellules ayant les caractéristiques des CSI [15].
À l’inverse, Hairless jouerait un rôle de senseur afin de bloquer la transcription basale des gènes cibles de la voie Notch dans la CSI, permettant ainsi de maintenir son identité [15]. Ce répresseur est également connu pour son rôle dans le recrutement de facteurs de remodelage de la chromatine [19], un processus important pour le maintien de l’homéostasie de l’intestin. En effet, le gène scrawny, codant pour une désubiquitinase de l’histone H2B, serait requis pour réprimer l’expression des gènes cibles de la voie Notch et permettre ainsi le maintien des CSI [20].
Différents niveaux d’activité de la voie Notch pour différents destins cellulaires
Le double rôle de la voie Notch, à la fois dans l’engagement de la CSI dans la voie de différenciation et dans le choix entre les destins entéroendocrine et entérocyte, pose la question des mécanismes de régulation de ces étapes. Plusieurs données génétiques indiquent que différents niveaux d’activation de la voie Notch seraient requis pour chacune des deux étapes de différenciation cellulaire au sein du lignage intestinal (Figure 2B).
Dans des cellules de mammifères en culture, il a été montré que l’activité du récepteur Notch peut être stimulée par sa fucosylation [21]. Dans l’intestin de drosophile, la perte d’activité des enzymes nécessaires à la fucosylation entraîne une accumulation de CSI sans perturber la différenciation terminale, c’est-à-dire la production de cellules entéroendocrines et entérocytes [22]. Des effets similaires sont observés dans le cas d’une mutation du gène rumi qui code pour une glycosyltransférase modifiant le domaine extracellulaire du récepteur Notch [23]. Grâce à ce mutant thermosensible, il est possible de reproduire les phénotypes associés aux différents niveaux d’activation de la voie Notch en fonction de la température. Une température élevée inhibe la voie Notch et entraîne l’accumulation de CSI et l’absence d’entérocytes. À l’inverse, une température intermédiaire permet une faible activation de la voie qui se traduit par une accumulation de CSI sans perturber la différenciation terminale des entéroblastes. En d’autres termes, une forte activation de la voie Notch serait nécessaire à la différenciation des CSI en entéroblastes, et un niveau plus faible d’activation serait suffisant pour déclencher la différenciation terminale (Figure 2B) [22]. Il a également été proposé que différents niveaux d’activation de la voie Notch participent au choix entre les destins entéroendocrine et entérocyte : un niveau plus élevé d’activité est requis pour la formation d’un entérocyte par rapport à celle d’une cellule entéroendocrine (Figure 2B) [14].
Les facteurs en aval de la voie Notch nécessaires pour le choix d’orientation entre cellules entéroendocrines et entérocytes n’ont pas été identifiés. Toutefois, d’autres voies de signalisation sont requises à cette étape de différenciation. Ainsi, la voie JAK (Janus kinase)/STAT (signal transducer and activator of transcription) participe à la formation des cellules différenciées, comme l’indique l’accumulation de cellules ayant les caractéristiques des entéroblastes en contexte mutant [24–26]. La voie EGF-R (epidermal growth factor receptor) est, quant à elle, requise dans les entérocytes pour la morphogenèse de l’intestin [27]. Les relations entre ces voies de signalisation et la voie Notch restent cependant à déterminer.
Un certain nombre de points restent à éclaircir concernant le rôle de Notch dans le lignage intestinal. Tout d’abord, les mécanismes à l’origine de l’activation asymétrique de la voie Notch dans une seule des deux cellules filles sont mal compris. Il a également été proposé que les CSI se divisent de manière symétrique [28–30]. Les deux nouvelles cellules souches produites n’activeraient pas la voie Notch. Les mécanismes de répression de la voie Notch dans le cas de ces divisions symétriques restent inconnus.
Le maintien de l’intégrité tissulaire
L’intestin est un organe en contact permanent avec le milieu extérieur. Ainsi, en plus du renouvellement naturel du tissu, l’intestin doit faire face à des agressions externes comme les stress physiques (lésions), chimiques (stress oxydatif) ou encore infectieux. Pour restaurer l’intégrité du tissu et assurer un état physiologique fonctionnel de l’organe, les CSI vont être activement sollicitées. Une régulation fine de leur capacité proliférative est alors indispensable et est assurée par un réseau complexe de voies de signalisation que nous allons détailler (Figure 3). Ce réseau entraîne principalement l’activation de deux voies centrales : les voies JAK/STAT et du récepteur de l’EGF (EGF-R).
 |
Figure 3. Contrôle de la prolifération des CSI à l’état d’homéostasie tissulaire et en réponse à un stress. A. Dans des conditions d’homéostasie tissulaire, la prolifération des cellules souches intestinales (CSI) est stimulée par les voies JAK/STAT, du récepteur à l’EGF (EGF-R), du récepteur à Insuline (InR) et par la voie Wingless (Wg). Ces voies sont, entre autres, activées dans les CSI par les ligands Vein, Wg et le peptide à insuline DiLP3, exprimés par les muscles viscéraux (MV), mais également par d’autres ligands de la voie de l’EGF-R exprimés par les CSI, les entéroblastes (EB) ou les entérocytes (EC). La voie de l’InR serait davantage impliquée dans l’adaptation du tissu face aux variations des apports nutritionnels. B. Lors d’une infection bactérienne ou au cours du vieillissement tissulaire, plusieurs voies de signalisation stimulent la prolifération des cellules souches. Au cours de cette réponse, une augmentation intracellulaire de ROS (reactive oxygen species) intervient, suivie de l’activation de la voie JNK (Jun N-terminal kinase) dans les EC. Après un dommage cellulaire, la voie Hippo est inhibée, ce qui entraîne une augmentation de la prolifération des CSI. Ainsi, l’activation de JNK, l’inhibition de Hippo, et l’intervention d’autres signaux mal connus impliqués dans la réponse aux dommages, stimulent la voie JAK/STAT par l’intermédiaire du ligand Upd3, et celle de l’EGF-R via l’expression musculaire du ligand Vein, dans les CSI. |
De nombreuses stratégies assurent une détection des dommages tissulaires au sein de l’intestin. La voie Hippo agit comme un senseur de l’intégrité tissulaire [56]. En absence de stress, elle restreint la capacité proliférative des CSI. À l’inverse, une perte de l’intégrité épithéliale provoque l’inactivation de cette voie, ce qui permet la stimulation de la prolifération des CSI [31–34]. Lors d’une activation constitutive de la voie Hippo, le tissu est incapable de répondre aux stress environnementaux (stress réplicatif, infection bactérienne, destruction de la lame basale), comme l’illustre l’absence de prolifération des cellules souches.
Dans le cas d’une mort cellulaire importante, ou encore d’un stress oxydatif (caractérisé par une forte concentration de dérivés réactifs de l’oxygène ROS [reactive oxygen species]), la réparation tissulaire est également associée à une activation de la voie de signalisation JNK (Jun N-terminal kinase) dans les entérocytes [26, 35, 36]. La voie JNK stimule la sécrétion du ligand Unpaired 3 (Upd 3) par les entérocytes et des ligands de la voie EGF-R par les cellules épithéliales et musculaires, ce qui induit l’activation des voies JAK/STAT et de l’EGF-R dans les CSI (Figure 3B) [27, 31–34, 37–40].
Dans le cas d’une infection bactérienne, la réponse au stress se fait en deux étapes. La réponse immunitaire va, dans un premier temps, se mettre en place. Elle se traduit par une rapide production de ROS qui, dans un second temps, stimulent la prolifération des CSI [39–41].
Les relations d’épistasie qui lient ces différentes voies de signalisation restent cependant mal connues. Certaines données placeraient la voie de l’EGF-R en aval de la voie JAK/STAT [26], alors que d’autres indiquent une action parallèle de ces deux voies [27, 42]. Les voies Hippo et des JNK semblent, quant à elles, agir en amont en modulant l’expression des ligands des voies JAK/STAT et de l’EGF-R [27, 31–34, 37–39]. D’autres voies de signalisation, comme l’activation de la kinase Dp38p, sont également impliquées dans la réponse au stress [43, 44].
En résumé, la mise en place des mécanismes de réparation tissulaire se fait par le biais de différentes voies de détection des dommages (JNK, Hippo) qui vont générer l’activation de voies prolifératives secondaires (JAK-STAT et EGF-R).
Le renouvellement naturel du tissu
En plus d’être impliquées dans la réparation tissulaire, les voies JAK/STAT [24, 26, 36, 39, 45–47] et de l’EGF-R [27, 37, 38, 42] exercent un rôle stimulateur sur la prolifération des CSI au cours du renouvellement naturel de l’épithélium, tout en assurant le maintien des CSI (Figure 3A). Autrement dit, l’absence de ces voies induirait une perte des CSI au sein du tissu [42]. L’étude de mutants affectant la transduction des signaux de la voie Wingless/Wnt, comme une perte de fonction des gènes codant pour le récepteur Frizzled, le facteur de transcription Armadillo/β-caténine ou encore la protéine effectrice Dishevelled, implique cette voie dans le maintien et la prolifération des CSI en absence de stress extrinsèques [48, 49]. De plus, une perte de fonction du gène suppresseur de tumeur apc (adenomatous polyposis coli), inhibiteur de la voie Wingless, est à l’origine d’une hyperplasie de l’épithélium. Ainsi, une activation constitutive de cette voie dans les CSI stimule leur prolifération sans affecter la différenciation des CSI nouvellement formées en entérocytes et cellules entéroendocrines [48, 49].
Afin de s’adapter aux variations environnementales telles que les fluctuations de nourriture, l’épithélium peut modifier sa croissance et la capacité proliférative des CSI. En effet, le niveau d’expression local du ligand DiLP3 (Drosophila Insulin-like peptide 3) sécrété par les muscles viscéraux (Figure 3A) peut influencer la prolifération des CSI par activation du récepteur à l’insuline [29, 50–52]. Cette augmentation du taux de division des CSI s’accompagnerait d’une tendance des CSI à se diviser symétriquement. Ces effets permettraient de réguler la croissance de l’intestin en augmentant le nombre total de cellules de cet organe afin d’assurer une meilleure adaptabilité du tissu face aux variations en apport nutritionnel (jeûne ou nutrition) [29, 51, 52]. Ainsi, la voie de l’insuline agirait d’avantage comme un signal permissif d’activation de la capacité proliférative des CSI en réponse à un apport de nutriments [53]. Ces analyses ouvrent un nouveau champ d’investigation sur le rôle de la voie du récepteur à l’insuline dans la modification du mode de division des CSI (asymétrique versus symétrique) qui est à ce jour peu connu.
Quiescence cellulaire et vieillissement
Le contrôle de l’état prolifératif des CSI en conditions d’homéostasie ou de réparation tissulaire est dépendant de la régulation de l’équilibre intracellulaire du système d’oxydoréduction. Cet équilibre serait nécessaire au maintien du réservoir de cellules souches quiescentes dans le tissu. Une faible quantité de ROS dans les CSI aurait un rôle limitant dans la division des CSI, empêchant ainsi toute prolifération excessive [41]. À l’inverse, une augmentation de ces ROS semble être impliquée dans la transition des CSI d’un état de quiescence à un état actif [41]. Les mécanismes moléculaires qui permettent de discriminer une CSI quiescente d’une CSI active ne sont cependant pas connus, ni ceux qui sont impliqués dans la transition entre ces deux états.
Le vieillissement tissulaire est caractérisé par une organisation anormale de l’épithélium, une augmentation dérégulée de la prolifération des CSI, une accumulation de cellules entéroblastes, ou encore par la présence d’entérocytes exprimant le marqueur des CSI : le ligand Delta. Cette désorganisation tissulaire s’accompagne d’une concentration cellulaire excessive de ROS et d’une suractivation de la voie de signalisation des JNK, toutes deux à l’origine de l’augmentation anormale de la prolifération des CSI, comme le suggère l’analogie de ces effets avec ceux que provoque une exposition à des agents oxydants comme le paraquat [35, 43, 54]. Ainsi, l’augmentation du nombre de CSI au cours du vieillissement pourrait être liée à une activation quasi constante de CSI quiescentes.
Une niche intestinale ?
L’épithélium intestinal est entouré par des muscles viscéraux produisant les contractions musculaires nécessaires à la digestion. Ceux-ci sécrètent également des facteurs extracellulaires mitogènes, tels que les ligands des voies de signalisation Wingless, de l’EGF-R, et de l’insuline [27, 29, 37, 38, 42, 47, 48, 55], à l’origine d’un environnement extracellulaire propice à la prolifération des CSI (Figure 3). L’expression de facteurs mitogènes n’est cependant pas exclusive aux muscles viscéraux. En effet, des ligands du récepteur de l’EGF et du récepteur de la voie JAK/STAT sont aussi exprimés au niveau des CSI, des entéroblastes et dans les entérocytes [26, 27, 37, 39, 42]. Par exemple, la sécrétion du ligand Unpaired 3 par les entérocytes au cours de la réparation tissulaire stimule l’expression des ligands des voies JAK/STAT et de l’EGF-R dans les CSI. En plus d’une stimulation de la prolifération des CSI par les entérocytes, une boucle homéostatique de régulation de l’activation des CSI est mise en place. Elle est à l’origine d’une stimulation autocrine des CSI, et donc d’une amplification du signal prolifératif. En résumé, l’environnement cellulaire participe au contrôle de la prolifération des CSI en produisant des signaux extracellulaires mitogènes.
Ce regroupement localisé de signaux, indépendant d’une structure physique définie, constitue un microenvironnement favorable à la régulation de la prolifération des CSI. Cependant, la question de l’existence d’une niche, c’est-à-dire d’une entité physique nécessaire au maintien de l’état « souche » au sein du tissu intestinal de drosophile est toujours en débat. Au sein de l’épithélium, les CSI ont une position basale et sont donc proches des muscles viscéraux [14]. Ainsi, ces derniers pourraient jouer ce rôle de niche pour les CSI. Toutefois, l’absence de fonction claire de ces muscles dans le maintien des CSI ne permet pas de conclure définitivement.
L’utilisation de l’intestin de drosophile comme modèle d’étude offre une meilleure compréhension des lois biologiques qui régulent les cellules souches au sein d’un tissu à renouvellement rapide. Cependant, de nombreuses questions restent à élucider. Une poursuite de l’effort de recherche sur ce modèle permettra d’améliorer nos connaissances dans ce domaine.
Liens d’intérêt
Les auteurs déclarent n’avoir aucun lien d’intérêt concernant les données publiées dans cet article.
Références
- Jones DL, Rando TA. Emerging models and paradigms for stem cell ageing. Nat Cell Biol 2011 ; 13 : 506–512. [CrossRef] [PubMed] [Google Scholar]
- Clevers H. The cancer stem cell: premises, promises and challenges. Nat Med 2011 ; 17 : 313–319. [CrossRef] [PubMed] [Google Scholar]
- Visvader JE, Lindeman GJ. Cancer stem cells in solid tumours: accumulating evidence and unresolved questions. Nat Rev Cancer 2008 ; 8 : 755–768. [CrossRef] [PubMed] [Google Scholar]
- Kiskinis E, Eggan K. Progress toward the clinical application of patient-specific pluripotent stem cells. J Clin Invest 2010 ; 120 : 51–59. [CrossRef] [PubMed] [Google Scholar]
- Jagut M, Huynh JR. Régulation des cellules souches de la lignée germinale : la niche s’agrandit chez la drosophile. Med Sci (Paris) 2007 ; 23 : 611–618. [CrossRef] [EDP Sciences] [PubMed] [Google Scholar]
- Losick VP, Morris LX, Fox DT, Spradling A. Drosophila stem cell niches: a decade of discovery suggests a unified view of stem cell regulation. Dev Cell 2011 ; 21 : 159–171. [CrossRef] [PubMed] [Google Scholar]
- Knoblich JA. Asymmetric cell division: recent developments and their implications for tumour biology. Nat Rev Mol Cell Biol 2010 ; 11 : 849–860. [CrossRef] [PubMed] [Google Scholar]
- Micchelli CA, Perrimon N. Evidence that stem cells reside in the adult Drosophila midgut epithelium. Nature 2006 ; 439 : 475–479. [CrossRef] [PubMed] [Google Scholar]
- Ohlstein B, Spradling A. The adult Drosophila posterior midgut is maintained by pluripotent stem cells. Nature 2006 ; 439 : 470–474. [CrossRef] [PubMed] [Google Scholar]
- Fre S, Bardin A, Robine S, Louvard D. Notch signaling in intestinal homeostasis across species: the cases of Drosophila, zebrafish and the mouse. Exp Cell Res 2011 ; 317 : 2740–2747. [CrossRef] [PubMed] [Google Scholar]
- Casali A, Batlle E. Intestinal stem cells in mammals and Drosophila. Cell Stem Cell 2009 ; 4 : 124–127. [CrossRef] [PubMed] [Google Scholar]
- Jiang H, Edgar BA. Intestinal stem cell function in Drosophila and mice. Curr Opin Genet Dev 2012 ; 22 : 354–360. [CrossRef] [PubMed] [Google Scholar]
- Brou C, Logeat F. Endocytose et voie de signalisation Notch. Med Sci (Paris) 2006 ; 22 : 685–688. [CrossRef] [EDP Sciences] [PubMed] [Google Scholar]
- Ohlstein B, Spradling A. Multipotent Drosophila intestinal stem cells specify daughter cell fates by differential notch signaling. Science 2007 ; 315 : 988–992. [CrossRef] [PubMed] [Google Scholar]
- Bardin AJ, Perdigoto CN, Southall TD, et al. Transcriptional control of stem cell maintenance in the Drosophila intestine. Development 2010 ; 137 : 715–724. [CrossRef] [PubMed] [Google Scholar]
- Kopan R, Ilagan MX. The canonical Notch signaling pathway: unfolding the activation mechanism. Cell 2009 ; 137 : 216–233. [CrossRef] [PubMed] [Google Scholar]
- Bray SJ. Notch signalling: a simple pathway becomes complex. Nat Rev Mol Cell Biol 2006 ; 7 : 678–689. [CrossRef] [PubMed] [Google Scholar]
- Kageyama R, Ohtsuka T, Kobayashi T. The Hes gene family: repressors and oscillators that orchestrate embryogenesis. Development 2007 ; 134 : 1243–1251. [CrossRef] [PubMed] [Google Scholar]
- Goodfellow H, Krejci A, Moshkin Y, et al. Gene-specific targeting of the histone chaperone asf1 to mediate silencing. Dev Cell 2007 ; 13 : 593–600. [CrossRef] [PubMed] [Google Scholar]
- Buszczak M, Paterno S, Spradling AC. Drosophila stem cells share a common requirement for the histone H2B ubiquitin protease scrawny. Science 2009 ; 323 : 248–251. [CrossRef] [PubMed] [Google Scholar]
- Stahl M, Uemura K, Ge C, et al. Roles of Pofut1 and O-fucose in mammalian Notch signaling. J Biol Chem 2008 ; 283 : 13638–13651. [CrossRef] [PubMed] [Google Scholar]
- Perdigoto CN, Schweisguth F, Bardin AJ. Distinct levels of Notch activity for commitment and terminal differentiation of stem cells in the adult fly intestine. Development 2011 ; 138 : 4585–4595. [CrossRef] [PubMed] [Google Scholar]
- Acar M, Jafar-Nejad H, Takeuchi H, et al. Rumi is a CAP10 domain glycosyltransferase that modifies Notch and is required for Notch signaling. Cell 2008 ; 132 : 247–258. [CrossRef] [PubMed] [Google Scholar]
- Beebe K, Lee WC, Micchelli CA. JAK/STAT signaling coordinates stem cell proliferation and multilineage differentiation in the Drosophila intestinal stem cell lineage. Dev Biol 2010 ; 338 : 28–37. [CrossRef] [PubMed] [Google Scholar]
- Buchon N, Poidevin M, Kwon HM, et al. A single modular serine protease integrates signals from pattern-recognition receptors upstream of the Drosophila Toll pathway. Proc Natl Acad Sci USA 2009 ; 106 : 12442–12447. [CrossRef] [Google Scholar]
- Jiang H, Patel PH, Kohlmaier A, et al. Cytokine/Jak/Stat signaling mediates regeneration and homeostasis in the Drosophila midgut. Cell 2009 ; 137 : 1343–1355. [CrossRef] [PubMed] [Google Scholar]
- Buchon N, Broderick NA, Kuraishi T, Lemaitre B. Drosophila EGFR pathway coordinates stem cell proliferation, gut remodeling following infection. BMC Biol, 2010 ; 8 : 152. [CrossRef] [PubMed] [Google Scholar]
- McLeod CJ, Wang L, Wong C, Jones DL. Stem cell dynamics in response to nutrient availability. Curr Biol 2010 ; 20 : 2100–2105. [CrossRef] [PubMed] [Google Scholar]
- O’Brien LE, Soliman SS, Li X, Bilder D. Altered modes of stem cell division drive adaptive intestinal growth. Cell 2011 ; 147 : 603–614. [CrossRef] [PubMed] [Google Scholar]
- de Navascues J, Perdigoto CN, Bian Y, et al. Drosophila midgut homeostasis involves neutral competition between symmetrically dividing intestinal stem cells. EMBO J 2012 ; 31 : 2473–2485. [CrossRef] [PubMed] [Google Scholar]
- Karpowicz P, Perez J, Perrimon N. The Hippo tumor suppressor pathway regulates intestinal stem cell regeneration. Development 2010 ; 137 : 4135–4145. [CrossRef] [PubMed] [Google Scholar]
- Shaw RL, Kohlmaier A, Polesello C, et al. The Hippo pathway regulates intestinal stem cell proliferation during Drosophila adult midgut regeneration. Development 2010 ; 137 : 4147–4158. [CrossRef] [PubMed] [Google Scholar]
- Staley BK, Irvine KD. Warts and Yorkie mediate intestinal regeneration by influencing stem cell proliferation. Curr Biol 2010 ; 20 : 1580–1587. [CrossRef] [PubMed] [Google Scholar]
- Ren F, Wang B, Yue T, et al. Hippo signaling regulates Drosophila intestine stem cell proliferation through multiple pathways. Proc Natl Acad Sci USA 2010 ; 107 : 21064–21069. [CrossRef] [Google Scholar]
- Biteau B, Hochmuth CE, Jasper H. JNK activity in somatic stem cells causes loss of tissue homeostasis in the aging Drosophila gut. Cell Stem Cell 2008 ; 3 : 442–455. [CrossRef] [PubMed] [Google Scholar]
- Buchon N, Broderick NA, Poidevin M, et al. Drosophila intestinal response to bacterial infection: activation of host defense and stem cell proliferation. Cell Host Microbe 2009 ; 5 : 200–211. [CrossRef] [PubMed] [Google Scholar]
- Jiang H, Grenley MO, Bravo MJ, et al. EGFR/Ras/MAPK signaling mediates adult midgut epithelial homeostasis and regeneration in Drosophila. Cell Stem Cell 2011 ; 8 : 84–95. [CrossRef] [PubMed] [Google Scholar]
- Biteau B, Jasper H. EGF signaling regulates the proliferation of intestinal stem cells in Drosophila. Development 2011 ; 138 : 1045–1055. [CrossRef] [PubMed] [Google Scholar]
- Buchon N, Broderick NA, Chakrabarti S, Lemaitre B. Invasive and indigenous microbiota impact intestinal stem cell activity through multiple pathways in Drosophila. Genes Dev 2009 ; 23 : 2333–2344. [CrossRef] [PubMed] [Google Scholar]
- Ha EM, Oh CT, Bae YS, Lee WJ. A direct role for dual oxidase in Drosophila gut immunity. Science 2005 ; 310 : 847–850. [CrossRef] [PubMed] [Google Scholar]
- Hochmuth CE, Biteau B, Bohmann D, Jasper H. Redox regulation by Keap1 and Nrf2 controls intestinal stem cell proliferation in Drosophila. Cell Stem Cell 2011 ; 8 : 188–199. [CrossRef] [PubMed] [Google Scholar]
- Xu N, Wang SQ, Tan D, et al. EGFR, Wingless and JAK/STAT signaling cooperatively maintain Drosophila intestinal stem cells. Dev Biol 2011 ; 354 : 31–43. [CrossRef] [PubMed] [Google Scholar]
- Choi NH, Kim JG, Yang DJ, et al. Age-related changes in Drosophila midgut are associated with PVF2, a PDGF/VEGF-like growth factor. Aging Cell 2008 ; 7 : 318–334. [CrossRef] [PubMed] [Google Scholar]
- Park JS, Kim YS, Yoo MA. The role of p38b MAPK in age-related modulation of intestinal stem cell proliferation and differentiation in Drosophila. Aging (Albany NY) 2009 ; 1 : 637–651. [PubMed] [Google Scholar]
- Cronin SJ, Nehme NT, Limmer S, et al. Genome-wide RNAi screen identifies genes involved in intestinal pathogenic bacterial infection. Science 2009 ; 325 : 340–343. [CrossRef] [PubMed] [Google Scholar]
- Liu W, Singh SR, Hou SX. JAK-STAT is restrained by Notch to control cell proliferation of the Drosophila intestinal stem cells. J Cell Biochem 2010 ; 109 : 992–999. [PubMed] [Google Scholar]
- Lin G, Xu N, Xi R. Paracrine unpaired signaling through the JAK/STAT pathway controls self-renewal and lineage differentiation of Drosophila intestinal stem cells. J Mol Cell Biol 2009 ; 2 : 37–39. [CrossRef] [PubMed] [Google Scholar]
- Lin G, Xu N, Xi R. Paracrine Wingless signalling controls self-renewal of Drosophila intestinal stem cells. Nature 2008 ; 455 : 1119–1123. [CrossRef] [PubMed] [Google Scholar]
- Lee WC, Beebe K, Sudmeier L, Micchelli CA. Adenomatous polyposis coli regulates Drosophila intestinal stem cell proliferation. Development 2009 ; 136 : 2255–2264. [CrossRef] [PubMed] [Google Scholar]
- Biteau B, Karpac J, Supoyo S, et al. Lifespan extension by preserving proliferative homeostasis in Drosophila. PLoS Genet 2010 ; 6 : e1001159. [CrossRef] [PubMed] [Google Scholar]
- Choi NH, Lucchetta E, Ohlstein B. Nonautonomous regulation of Drosophila midgut stem cell proliferation by the insulin-signaling pathway. Proc Natl Acad Sci USA 2011 ; 108 : 18702–18707. [CrossRef] [Google Scholar]
- Wang L, McLeod CJ, Jones DL. Regulation of adult stem cell behavior by nutrient signaling. Cell Cycle 2011 ; 10 : 2628–2634. [CrossRef] [PubMed] [Google Scholar]
- Amcheslavsky A, Jiang J, Ip YT. Tissue damage-induced intestinal stem cell division in Drosophila. Cell Stem Cell 2009 ; 4 : 49–61. [CrossRef] [PubMed] [Google Scholar]
- Choi YJ, Hwang MS, Park JS, et al. Age-related upregulation of Drosophila caudal gene via NF-kappaB in the adult posterior midgut. Biochim Biophys Acta 2008 ; 1780 : 1093–1100. [CrossRef] [PubMed] [Google Scholar]
- Veenstra JA, Agricola HJ, Sellami A. Regulatory peptides in fruit fly midgut. Cell Tissue Res 2008 ; 334 : 499–516. [CrossRef] [PubMed] [Google Scholar]
- Bardet PL. La voie Hippo contrôle la croissance des organes au cours du développement. Med Sci (Paris) 2009 ; 25 : 253–257. [CrossRef] [EDP Sciences] [PubMed] [Google Scholar]
Liste des figures
|
Figure 1. Organisation structurelle et cellulaire de l’intestin moyen de drosophile. A. Le système digestif est connecté en son extrémité antérieure à la bouche, aux glandes salivaires (non représentées) et au jabot. Il se prolonge jusqu’à l’intestin antérieur composé, entre autres, de l’œsophage. Ce dernier aurait une fonction similaire à celle de l’estomac. L’appareil digestif est également composé d’un intestin moyen et d’un intestin postérieur. L’intestin moyen de drosophile est segmenté en trois parties majeures : une région antérieure (rouge), une région intermédiaire (vert) qui inclut une zone composée de cellules Cu2+ au pH très acide (pH < 3), et une région postérieure (bleu). B. L’épithélium intestinal est entouré de cellules musculaires viscérales (MV), et est composé de cellules différenciées : les cellules entéroendocrines (ee, cellules sécrétrices) et les entérocytes (EC, cellules absorbantes), et de cellules progénitrices : les cellules souches intestinales (CSI) et les entéroblastes (EB, cellules post-mitotiques). C. Le lignage intestinal cellulaire est simple. Les cellules souches intestinales sont les seules cellules capables de se diviser et la majorité d’entre elles génèrent, après division, une nouvelle cellule souche et un entéroblaste. Cet entéroblaste a la capacité de poursuivre sa différenciation pour devenir soit une cellule entéroendocrine (ee), soit un entérocyte (EC). |
| Dans le texte | |
|
Figure 2. Modèle de régulation de la voie Notch dans le lignage intestinal. A. En l’absence d’activation de la voie Notch, Hairless permettrait le maintien des cellules souches (CSI) en réprimant l’expression des gènes cibles de la voie. L’activité du facteur de transcription Daughterless (Da) serait ainsi maintenue entraînant le renouvellement des CSI et le maintien de l’expression de Delta. L’activation de la voie Notch par le ligand Delta dans la cellule voisine entraîne le clivage du récepteur Notch et la translocation de son domaine intracellulaire (NICD) dans le noyau. Le complexe NICD/suppressor of hairless (Su[H]) active la transcription des gènes cibles de la voie Notch, parmi lesquels les gènes codant pour les membres du complexe enhancer of split (E[spl]-C). Le complexe E(spl)-C aurait comme fonction d’inhiber l’activité de Da, permettant ainsi la différenciation de la cellule en entéroblaste (EB). L’activité de la voie Notch est modulée par des modifications post-traductionnelles du récepteur Notch (fucosylation, glycosylation, en jaune). Aucune restriction de ces modifications aux EB n’a été rapportée jusqu’ici. RE : réticulum endoplasmique. B. Un niveau élevé d’activation de la voie Notch dépendant de la fucosylation du récepteur doit être atteint pour entraîner le basculement de la cellule souche dans la voie de différenciation (EB). Dans un deuxième temps, la différenciation terminale des entéroblastes en cellules entérocytes (EC) requiert un niveau plus faible d’activité de la voie Notch, indépendant de la fucosylation du récepteur. La relation temporelle entre ces événements n’est cependant pas caractérisée à l’heure actuelle. Il a également été proposé que différents niveaux d’activation de Notch participent au choix entre les destins des cellules entéroendocrines (ee) et des entérocytes (EC) : un niveau plus élevé serait requis pour former un entérocyte, alors qu’un faible niveau d’activation permettrait la différenciation de l’entéroblaste en cellule entéroendocrine. |
| Dans le texte | |
|
Figure 3. Contrôle de la prolifération des CSI à l’état d’homéostasie tissulaire et en réponse à un stress. A. Dans des conditions d’homéostasie tissulaire, la prolifération des cellules souches intestinales (CSI) est stimulée par les voies JAK/STAT, du récepteur à l’EGF (EGF-R), du récepteur à Insuline (InR) et par la voie Wingless (Wg). Ces voies sont, entre autres, activées dans les CSI par les ligands Vein, Wg et le peptide à insuline DiLP3, exprimés par les muscles viscéraux (MV), mais également par d’autres ligands de la voie de l’EGF-R exprimés par les CSI, les entéroblastes (EB) ou les entérocytes (EC). La voie de l’InR serait davantage impliquée dans l’adaptation du tissu face aux variations des apports nutritionnels. B. Lors d’une infection bactérienne ou au cours du vieillissement tissulaire, plusieurs voies de signalisation stimulent la prolifération des cellules souches. Au cours de cette réponse, une augmentation intracellulaire de ROS (reactive oxygen species) intervient, suivie de l’activation de la voie JNK (Jun N-terminal kinase) dans les EC. Après un dommage cellulaire, la voie Hippo est inhibée, ce qui entraîne une augmentation de la prolifération des CSI. Ainsi, l’activation de JNK, l’inhibition de Hippo, et l’intervention d’autres signaux mal connus impliqués dans la réponse aux dommages, stimulent la voie JAK/STAT par l’intermédiaire du ligand Upd3, et celle de l’EGF-R via l’expression musculaire du ligand Vein, dans les CSI. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.