")
")
| Issue |
Med Sci (Paris)
Volume 24, Number 10, Octobre 2008
|
|
|---|---|---|
| Page(s) | 853 - 858 | |
| Section | M/S revues | |
| DOI | https://doi.org/10.1051/medsci/20082410853 | |
| Published online | 15 October 2008 | |
Pathogénie du syndrome néphrotique à lesions glomérulaires minimes
Minimal change nephrotic syndrome: new insights into disease pathogenesis
Inserm U 841, Institut Mondor de Recherche Biomédicale (IMRB), Département « génétique », Équipe « 21 » ; AP-HP, Groupe hospitalier Henri Mondor-Albert Chenevier, Service de Néphrologie, 94010 Créteil, France
*
This email address is being protected from spambots. You need JavaScript enabled to view it.
Résumé
Le syndrome néphrotique idiopathique (SNI) est une néphropathie glomérulaire définie par une protéinurie massive associée à une hypoalbuminémie, sans lésions inflammatoires rénales ni dépôts de complexes immuns circulants. Le SNI à lésions glomérulaires minimes (SNLGM) représente la forme la plus fréquente des syndromes néphrotiques et se caractérise par des perturbations du système immunitaire et une atteinte rénale podocytaire dont le mécanisme reste obscur. La phase active de la maladie est associée à une augmentation du nombre de lymphocytes T et B et à une production accrue de cytokines. Les principales perturbations lymphocytaires qui semblent caractéristiques du SNLGM sont l’inhibition de l’hypersensibilité de type retardé, les anomalies de la commutation isotypique et une polarisation lymphocytaire T qui ne correspond ni à une voie de type Th1 ou Th2 classique et qui résulte d’une activation transcriptionnelle anormale. Le lien moléculaire entre l’atteinte immunologique et l’atteinte podocytaire reste non élucidé.
Abstract
Idiopathic nephrotic syndrome is the most frequent glomerular disease in children. While genetic analyses have provided new insights into disease pathogenesis through the discovery of several podocyte genes mutated in distinct forms of inherited nephrotic syndrome, the molecular bases of minimal change nephrotic syndrome (MCNS) and focal and segmental glomerulosclerosis (FSGS) with relapse remain unclear. Although immune cell disorders, which may involve both innate and adaptive immunity, appear to play a role in the pathogenesis of steroid sensitive MCNS, the mechanisms by which they induce podocyte dysfunction remain unresolved. It was postulated that podocyte injury results from a circulating factor secreted by abnormal T cells, but the possibility that bipolarity of the disease results from a functional disorder shared by both cell systems is not excluded. MCNS relapses are associated with an activation of the immune system, including an expansion of T and B cell compartments and production of growth factors as well as many cytokines. Dysfunction of T cells is supported by three main findings: (1) inhibition of a type III hypersensitivity reaction ; (2) defects in immunoglobulin switch ; (3) unclassical T helper polarization resulting from transcriptional interference between Th1 and Th2 transcriptional factors.
© 2008 médecine/sciences - Inserm / SRMS
Le syndrome néphrotique à lésions glomérulaires minimes (SNLGM) est une entité anatomo-clinique définie par une protéinurie sélective massive (> 3g/jour) et une hypoalbuminémie (< 30 g/l) sans lésions glomérulaires visibles en microscopie optique et sans dépôts d’immunoglobulines ou de compléments en immunofluorescence. Le SNLGM représente la forme la plus fréquente de syndrome néphrotique idiopathique (SNI) qui inclut au moins deux autres entités histologiques caractérisées par des lésions de hyalinose segmentaire et focale (HSF) ou de prolifération mésangiale. Le SNI représente près de 90 % des néphropathies d’origine glomérulaire de l’enfant et 15 à 20 % de celles de l’adulte.
La réponse au traitement corticoïde permet de distinguer les formes sensibles qui sont d’un bon pronostic des formes résistantes qui peuvent évoluer vers la glomérulosclérose et l’insuffisance rénale chronique, en l’absence de réponse aux traitements prescrits habituellement en deuxième ligne (anti-calcineurine, cyclophosphamide). On considère que les formes résistantes aux divers traitements mais qui ne récidivent pas après la transplantation rénale, sont causées par des anomalies intrinsèques de gènes (mutations ou délétions) dont les produits sont impliqués dans le fonctionnement de la barrière de filtration glomérulaire. En revanche, les formes corticosensibles seraient liées à des altérations du système immunitaire.
Bien que les formes corticosensibles jouissent d’un pronostic rénal favorable à long terme, les rechutes sont fréquentes lorsque les doses de corticoïdes sont réduites ou après l’arrêt du traitement, nécessitant chez certains patients l’introduction de drogues immunosuppressives comme les anti-calcineurine ou les agents alkylants (cyclophosphamide, chloraminophène). La pathogénie des syndromes néphrotiques corticorésistants ne sera pas abordée dans cette minisynthèse.
Physiopathologie du syndrome néphrotique idiopathique
La fuite massive de protéines dans les urines résulte d’une hyperperméabilité de la barrière de filtration glomérulaire. Cette barrière est composée de trois types d’éléments : (1) l’endothélium fenestré du capillaire glomérulaire ; (2) la membrane basale glomérulaire (MBG) est une matrice extracellulaire acellulaire qui recouvre l’endothélium des capillaires glomérulaires et qui est composée essentiellement de Laminine 11 (α5, β2, γ1)1, de Perlecan (un protéoglycane à héparane sulphate), de collagène de type IV, de Nidogène et de protéoglycanes [1] ; (3) les cellules épithéliales viscérales différenciées appelées podocytes qui sont des cellules polarisées baignant dans le filtrat de l’espace de Bowman et ancrées à la membrane basale par des expansions membrano-cytoplasmiques appelées pédicelles (Figure 1). L’espace entre deux pédicelles adjacents forme une fente étroite de 40 nm de largeur appelée fente de filtration qui est recouverte sur son versant externe par une structure membranoïde hautement spécialisée appelée diaphragme de fente, lequel constitue l’ultime barrière de filtration qui s’oppose au passage de protéines dans la chambre urinaire [2, 3].
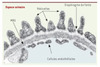 |
Figure 1. La barrière de filtration glomérulaire en microscopie électronique. MBG : membrane basale glomérulaire. |
L’étude de l’architecture moléculaire du podocyte permet de distinguer trois structures dynamiques qui contrôlent l’intégrité de la barrière de filtration (Figure 2) : (1) le diaphragme de fente qui est une jonction adhérente unique, formée par des homodimères de néphrine sur lesquels s’agencent d’autres molécules telles que FAT (un membre de la superfamille des cadhérines) et Neph 1 (une protéine des podocytes proche de la néphrine), la cadhérine P et la Densine ; (2) le domaine basal qui permet l’ancrage du podocyte dans la MBG par l’intermédiaire de molécules d’adhésion comme les intégrines α3β1 et les dystroglycanes ; (3) le cytosquelette des pédicelles qui interagit avec ces structures par l’intermédiaire de protéines exprimées dans les zones cellulaires proximales en particulier la podocine, CD2AP2 (CD2-associated protein) et l’ILK (integrin-linked kinase). L’étude de ces protéines a permis de montrer que tout processus qui interfère avec cette interconnexion dynamique est susceptible d’altérer le fonctionnement de la barrière de filtration et d’induire une protéinurie.
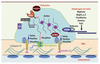 |
Figure 2. Vue schématique des pédicelles. ILK : integrin-linked kinase ; les hexagones verts symbolisent les intégrines ab ; MBG : membrane basale glomérulaire ; CD2AP : CD2 associated protein ; Glepp : glomerular epithelial protein. |
L’atteinte podocytaire dans le SNLGM résulte d’une altération des signaux cellulaires aboutissant à une perte de l’intégrité fonctionnelle qui est habituellement réversible sous traitement corticoïde et/ou immunosuppresseur.
Perturbations de la signalisation lymphocytaire dans le SNI corticosensible
L’étiologie du SNI d’origine dysimmunitaire reste une énigme. L’origine extra-rénale du SNI est suspectée sur les arguments cliniques et expérimentaux suivants : (1) la transplantation de reins de donneurs atteints de SNI chez des receveurs indemnes de cette maladie entraîne, en moins d’une semaine, une disparition complète du syndrome néphrotique lié à une LGM (lésions glomérulaires minimes) [4], ou une HSF (hyalinose segmentaire et focale) [5] ; (2) les échanges plasmatiques réalisés de façon exceptionnelle chez les patients en poussée résistant au traitement peuvent conduire à des rémissions ponctuelles suggérant que le facteur pathogénique est présent dans le sang périphérique [6, 7] ; (3) les surnageants de lymphocytes T périphériques isolés chez des patients atteints de SNI et activés in vitro, ainsi que le plasma prélevé au cours des rechutes ou lors des récidives après transplantation rénale, sont capables d’induire une protéinurie chez le rat [8–11] ; (4) la transmission materno-fœtale transitoire du syndrome néphrotique [12, 13].
Interférence transcriptionnelle dans les voies cytokiniques : l’exemple de l’IL-4
Les événements moléculaires qui s’enchaînent après l’agression initiale du podocyte sont inconnus mais ces événements altèrent le fonctionnement normal du cytosquelette podocytaire et modifient les interactions entre le podocyte et la matrice extracellulaire. Les études physiopathologiques des formes corticosensibles ont été quasi-exclusivement centrées sur les perturbations immunologiques. La Figure 3 résume les hypothèses pathogéniques actuelles. L’étude des fonctions lymphocytaires T au cours des phases aiguës du SNI a mis en évidence une inhibition de l’hypersensibilité de type retardée, une hyporéactivité cellulaire aux mitogènes et une polarisation T helper « contrariée », caractérisée par un profil cytokinique mixte associant une production accrue d’interleukines de type Th1 [IFN (interféron), interleukine (IL)-8] et de type Th2 (IL-13, IL-10). Ces résultats s’expliquent par l’activation concomitante et inappropriée au sein des mêmes cellules de voies transcriptionnelles antagonistes comme NF-κB, cmaf, AP1, T bet, ce qui semble être une caractéristique propre aux formes corticosensibles. La régulation de l’IL-4 illustre remarquablement bien la notion d’interférence transcriptionnelle au cours des SNI. Chez les patients en phase de poussée et en l’absence de tout terrain allergique préexistant, l’IL-4 n’est pas produite dans les lymphocytes T CD4+ et ce malgré l’induction du facteur transcriptonnel cmaf dont L’IL-4 est un gène cible majeur. Ce phénomène s’explique par une activation importante de la voie NF-κB qui s’oppose à la transcription du gène en bloquant l’accès de cmaf à son site de liaison sur le promoteur de l’IL-4 [14]. Chez les mêmes patients en phase de rémission, le niveau d’IL-4 s’élève significativement et est corrélé à l’inhibition de la voie NF-κB par les corticoïdes. Bien que les mécanismes moléculaires qui sous-tendent l’interférence transcriptionnelle restent à élucider, ces résultats suggèrent que la polarisation des lymphocytes T au cours des SNI est possiblement un événement secondaire. Des études récentes, basées sur l’étude du polymorphisme de la région variable du récepteur T n’ont pas détecté un recrutement oligoclonal d’une population lymphocytaire au cours des poussées [15].
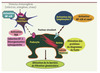 |
Figure 3. Mécanismes potentiels impliqués dans la pathogénie du syndrome néphrotique idiopathique. |
Les sous-populations lymphocytaires : rôle des lymphocytes T CD4+CD25+
L’analyse du phénotype des sous-populations lymphocytaires T périphériques, lors des rechutes, montre une expansion des lymphocytes T CD4+ qui expriment le marqueur CD25 (chaîne a du récepteur de l’IL-2) [16], ainsi que des lymphocytes T CD4+ et CD8+ qui expriment le marqueur CD45RO, caractéristique des lymphocytes T mémoires [17]. L’expression de l’antigène CD25 au cours des poussées peut refléter une activation des cellules T CD4+ ou bien traduire le recrutement d’une sous-population mineure (≈10 %) de lymphocytes T CD4+ CD25+, dotée de fonctions suppressives. Cette seconde éventualité est compatible avec certaines études fonctionnelles qui ont montré une diminution de l’immunité à médiation cellulaire appréciée par des tests cutanés d’hypersensibilité retardée (type IV dans la classification de Gell et Combs) et une réduction de la capacité des lymphocytes T à proliférer en présence de mitogènes [18, 19]. Cependant, le rôle spécifique des lymphocytes T suppresseurs dans le SNI reste à établir. Il est vraisemblable qu’une proportion majoritaire des cellules T CD4+ exprimant le marqueur CD25 correspond à des cellules T activées au cours des néphroses.
Altérations des populations lymphocytaires B
Les altérations lymphocytaires au cours du SNI ne sont pas limitées au seul compartiment T puisque au cours des poussées, l’on observe également une expansion significative de la population lymphocytaire B, mais celle-ci a été peu étudiée. Il semble que la différenciation B soit également affectée comme l’atteste la baisse fréquente des taux sériques des immunoglobulines. Celle-ci ne peut pas être expliquée par la seule fuite urinaire car elle n’affecte que certaines fractions d’IgG (IgG1, IgG2), ce qui suggère des anomalies de la coopération T/B au cours du SNI. Ces observations peuvent contribuer à la compréhension de l’effet bénéfique potentiel du traitement par des anticorps anti-CD20 au cours de certaines formes de SNI [20].
Dans certaines formes pédiatriques de SNI, les rechutes surviennent dans un délai très court suivant une infection, volontiers d’origine virale. La rapidité de survenue suggère un lien entre l’atteinte podocytaire et l’activation de l’immunité innée puisque à priori, l’immunité adaptative met deux à trois semaines pour être effective. Des données expérimentales réalisées chez la souris ont montré que l’injection de LPS (lipopolysaccharide), un antigène de la paroi bactérienne, induit l’expression podocytaire de novo de la molécule B7.1 et déclenche via sa liaison avec un Toll-like récepteur, TLR4, des cascades de signaux pour l’instant non identifiées, qui entraînent une désorganisation du cytosquelette et un syndrome néphrotique [21]. L’intérêt de ce modèle en pathologie humaine reste à démontrer, mais il ouvre toutefois de nouvelles pistes de recherches.
Altération de la signalisation podocytaire au cours du syndrome néphrotique idiopathique
Les données actuelles indiquent clairement que le diaphragme de fente n’est pas une structure statique mais une plateforme dynamique qui communique en continu avec le cytosquelette, via un recrutement séquentiel des protéines présentes dans les couches lipidiques de la membrane plasmique. Cette signalisation bidirectionnelle est cruciale pour le maintien de l’intégrité fonctionnelle et la survie du podocyte. Le déficit quantitatif ou qualitatif d’une protéine clé du diaphragme de fente comme la Néphrine interrompt la transduction des signaux vers l’intérieur du podocyte et entraîne une désorganisation du cytosquelette, une rétraction des pédicelles et un affaissement des podocytes.
Le rôle clé de Fyn et de la Néphrine
La Néphrine est le premier composant structural du diaphragme de fente, codée par le gène NPHS1 qui a été identifié par clonage positionnel à partir de l’étude de familles atteintes de syndrome néphrotique congénital de type Finlandais [22]. Il s’agit d’une protéine transmembranaire de 1 241 acides aminés avec un court domaine intracellulaire et un domaine extracellulaire comportant 8 domaines immunoglobuline-like (Ig-like) et un domaine transmembranaire de type fibronectine de type III. Chaque motif Ig-like est de type C2, caractéristique des protéines impliquées dans les interactions inter-cellulaires ou avec les composants de la matrice.
La Néphrine aurait un rôle de récepteur transmembranaire qui, à la suite de sa liaison avec son ligand, non encore identifié, va initier une cascade d’activations. Ce processus présente des similitudes avec celui qui met en jeu le récepteur clonotypique dans le lymphocyte T sauf que beaucoup d’acteurs moléculaires qui interviennent dans le lymphocyte ne sont pas exprimés dans le podocyte ou bien ne sont pas identifiés. La liaison de la Néphrine à son ligand entraîne un regroupement des complexes dans des microdomaines lipidiques (lipid rafts, ou radeaux lipidiques). Cet événement va activer la protéine Fyn, un membre de la famille Src kinase, qui phosphoryle plusieurs résidus tyrosine situés à l’extrémité amino-terminale de la Néphrine, au niveau desquels sont recrutées des protéine kinases et des protéines adaptatrices qui se lient ensuite à la Néphrine via leurs domaines SH2 [23–26].
La phosphorylation de la Néphrine par Fyn a deux conséquences essentielles (Figure 4) : (1) elle active la PI3K via l’interaction du domaine SH2 de la sous-unité régulatrice p85 avec le domaine cytoplasmique de la Néphrine, tandis que la sous-unité catalytique p110 génère des phospho-inositides capables de recruter et d’activer la sérine-thréonine kinase Akt, qui joue un rôle clé dans les processus de survie du podocyte ; (2) elle stabilise le complexe fonctionnel Néphrine-Podocine [26]. La déphosphorylation de la Néphrine provoque le détachement de la podocine et la liaison de la β-arrestine-2 à la Néphrine qui subit alors une endocytose suivie d’une dégradation, interrompant ainsi la voie de signalisation [27].
 |
Figure 4. Signalisation podocytaire médiée par la Néphrine. |
D’autres protéines du diaphragme de fente semblent jouer un rôle déterminant dans la transmission du signal vers le cytosquelette podocytaire. La protéine CD2 interagit directement avec p85 qui l’amène au contact du complexe néphrine-podocine, ce qui contribue à renforcer les signaux émanant de la plateforme de signalisation incluant les signaux anti-apoptotiques. En effet, Les podocytes de souris déficientes pour CD2AP sont caractérisés par une susceptibilité accrue à l’apoptose possiblement liée à la diminution de l’activité de Akt [28].
Autres cibles de Fyn
Outre son rôle sur l’activation de la Néphrine, Fyn interagit avec N-WASp (Wiskott Aldrich syndrome protein) qu’elle phosphoryle, entraînant le recrutement et l’ancrage de Nck aux microdomaines lipidiques [23, 24]. La famille des protéines adaptatrices Nck comprend Nck1 (Nckα) et Nck2 (Nckβ ou Grb4), qui possède chacune trois domaines SH3 et un domaine SH2 carboxy-terminal [29]. Nck se lie, via ses domaines SH3, à différentes protéines impliquées dans la régulation de l’actine telles que WASp et le complexe Arp2/3 [24], tandis que, via ses domaines SH2, Nck se lie aux phosphotyrosines de la Néphrine ce qui permet une connexion étroite entre la plate-forme de signalisation et l’organisation du cytosquelette d’actine. Le rôle de la phosphorylation de la Néphrine au cours de la morphogenèse podocytaire reste controversé comme en témoignent des données récentes qui montrent que les souris déficientes pour la néphrine ont un développement podocytaire normal bien que les podocytes n’expriment pas de diaphragme de fente [30].
Les résultats acquis in vitro et surtout in vivo suggèrent que Fyn joue un rôle primordial dans la régulation des signaux proximaux et dans le contrôle dynamique du cytosquelette. Toutefois, la régulation de Fyn dans le podocyte n’est pas connue, d’autant que les protéines qui contrôlent son activation dans le lymphocyte T sont peu ou pas exprimées par le podocyte normal (données personnelles).
L’étude de la signalisation podocytaire est actuellement à l’état embryonnaire. Les données disponibles font souvent l’objet de controverses et les résultats obtenus in vitro, voire dans des modèles animaux, peuvent ne pas reproduire ou ne pas rendre compte du phénotype observé chez l’homme. La situation est d’autant plus complexe que nos connaissances sur l’architecture moléculaire du diaphragme de fente ainsi que les interactions de celui-ci avec les régions proximales du podocytes ou avec le milieu environnant sont encore imparfaitement élucidées.
Chaque laminine est une glycoprotéine hétérotrimérique, dont les chaînes polypeptidiques individuelles sont liées par une liaison coiled-coil, produisant une molécule avec un bras long et 2-3 bras courts. La combinaison de plusieurs sous-unités α, β, et γ forme 15 laminines chez les mammifères.
Ainsi nommée car elle lie la partie intracytoplasmique de la molécule CD2 dans les lymphocytes T.
Références
- Tryggvason K, Patrakka J, Wartiovaara J. Hereditary proteinuria syndromes and mechanisms of proteinuria. N Engl J Med 2006; 354 : 1387–401. [Google Scholar]
- Pavenstadt H, Kriz W, Kretzler M. Cell biology of the glomerular podocyte. Physiol Rev 2003; 83 : 253–307. [Google Scholar]
- Deen WM. What determines glomerular capillary permeability ? J Clin Invest 2004; 114 : 1412–4. [Google Scholar]
- Ali AA, Wilson E, Moorhead JF, et al. Minimal-change glomerular nephritis. Normal kidneys in an abnormal environment ? Transplantation 1994; 58 : 849–52. [Google Scholar]
- Rea R, Smith C, Sandhu K, Kwan J, et al. Successful transplant of a kidney with focal segmental glomerulosclerosis. Nephrol Dial Transplant 2001; 16 : 416–7. [Google Scholar]
- Feld SM, Savin V, Nast CC, et al. Plasmapheresis in the treatment of steroid-resistant local segmental glomerulosclerosis in native kidneys. Am J Kidney Dis 1998; 32 : 230–7. [Google Scholar]
- Ginsburg DS, Dau P. Plasmapheresis in the treatment of steroid-resistant focal segmental glomerulosclerosis. Clin Nephrol 1997; 48 : 282–7. [Google Scholar]
- Lagrue G, Branellec A, Blanc C, et al. A vascular permeability factor in lymphocyte culture supernants from patients with nephrotic syndrome. II. Pharmacological and physicochemical properties. Biomedicine 1975; 23 : 73–5. [Google Scholar]
- Boulton J, JM, Tulloch I, et al. Changes in the glomerular capillary wall induced by lymphocyte products and serum of nephrotic patients. Clin Nephrol 1983; 20 : 72–7. [Google Scholar]
- Tanaka R, Yoshikawa N, Nakamura H, et al. Infusion of peripheral blood mononuclear cell products from nephrotic children increases albuminuria in rats. Nephron 1992; 60 : 35–41. [Google Scholar]
- Koyama A, Fujisaki M, Kobayashi M, et al. A glomerular permeability factor produced by human T cell hybridomas. Kidney Int 1991; 40 : 453–60. [Google Scholar]
- Lagrue G, Branellec A, Niaudet P, et al. Transmission of nephrotic syndrome to two neonates. Spontaneous regression. Presse Med 1991; 20 : 255–7. [Google Scholar]
- Kemper MJ, Wolf G, Muller-Wiefel DE. Transmission of glomerular permeability factor from a mother to her child. N Engl J Med 2001; 344 : 386–7. [Google Scholar]
- Valanciute A, le Gouvello S, Solhonne B, et al. NF-kappa B p65 antagonizes IL-4 induction by c-maf in minimal change nephrotic syndrome. J Immunol 2004; 172 : 688–98. [Google Scholar]
- Herve C, Le Berre L, Miqueu P, et al. Blood T-cell repertoire in idiopathic nephrotic syndrome recurrence following kidney transplantation. Am J Transplant 2006; 6 : 2144–51. [Google Scholar]
- Neuhaus TJ, Shah V, Callard, RE, Barratt TM. T-lymphocyte activation in steroid-sensitive nephrotic syndrome in childhood. Nephrol Dial Transplant 1995; 10 : 1348–52. [Google Scholar]
- Yan K, Nakahara K, Awa S, et al. The increase of memory T cell subsets in children with idiopathic nephrotic syndrome. Nephron 1998; 79 : 274–8. [Google Scholar]
- Fodor P, Saitua MT, Rodriguez E, et al. T-cell dysfunction in minimal-change nephrotic syndrome of childhood. Am J Dis Child 1982; 136 : 713–7. [Google Scholar]
- Matsumoto K, Osakabe K, Katayama H, et al. Defective cell-mediated immunity in lipoid nephrosis. Int Arch Allergy Appl Immunol 1984; 73 : 370–2. [Google Scholar]
- Francois H, Daugas E, Bensman A, Ronco P. Unexpected efficacy of rituximab in multirelapsing minimal change nephrotic syndrome in the adult: first case report and pathophysiological considerations. Am J Kidney Dis 2007; 49 : 158–61. [Google Scholar]
- Reiser J, von Gersdorff G, Loos M, et al Induction of B7-1 in podocytes is associated with nephrotic syndrome. J Clin Invest 2004; 113 : 1390–7. [Google Scholar]
- Kestila M, Lenkkeri U, Mannikko M, et al. Positionally cloned gene for a novel glomerular protein--nephrin--is mutated in congenital nephrotic syndrome. Mol Cell 1998; 1 : 575–82. [Google Scholar]
- Jones N, Blasutig IM, Eremina V, et al. Nck adaptor proteins link nephrin to the actin cytoskeleton of kidney podocytes. Nature 2006; 440 : 818–23. [Google Scholar]
- Verma R, Kovari I, Soofi A, et al. Nephrin ectodomain engagement results in Src kinase activation, nephrin phosphorylation, Nck recruitment, and actin polymerization. J Clin Invest 2006; 116 : 1346–59. [Google Scholar]
- Verma R, Wharram B, Kovari I, et al. Fyn binds to and phosphorylates the kidney slit diaphragm component Nephrin. J Biol Chem 2003; 278 : 20716–23. [Google Scholar]
- Li H, Lemay S, Aoudjit L, et al. SRC-family kinase Fyn phosphorylates the cytoplasmic domain of nephrin and modulates its interaction with podocin. J Am Soc Nephrol 2004; 15 : 3006–15. [Google Scholar]
- Quack I, Rump LC, Gerke P, et al. beta-Arrestin2 mediates nephrin endocytosis and impairs slit diaphragm integrity. Proc Natl Acad Sci USA 2006; 103 : 14110–5. [Google Scholar]
- Huber TB, Hartleben B, Kim J, et al. Nephrin and CD2AP associate with phosphoinositide 3-OH kinase and stimulate AKT-dependent signaling. Mol Cell Biol 2003; 23 : 4917–28. [Google Scholar]
- Chen M, She H, Davis E, et al. Identification of Nck family genes, chromosomal localization, expression, and signaling specificity. J Biol Chem 1998; 273 : 25171–8. [Google Scholar]
- Done SC, Takemoto M, He L, et al. Nephrin is involved in podocyte maturation but not survival during glomerular development. Kidney Int 2008; 73 : 697–704. [Google Scholar]
Liste des figures
|
Figure 1. La barrière de filtration glomérulaire en microscopie électronique. MBG : membrane basale glomérulaire. |
| Dans le texte | |
|
Figure 2. Vue schématique des pédicelles. ILK : integrin-linked kinase ; les hexagones verts symbolisent les intégrines ab ; MBG : membrane basale glomérulaire ; CD2AP : CD2 associated protein ; Glepp : glomerular epithelial protein. |
| Dans le texte | |
|
Figure 3. Mécanismes potentiels impliqués dans la pathogénie du syndrome néphrotique idiopathique. |
| Dans le texte | |
|
Figure 4. Signalisation podocytaire médiée par la Néphrine. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.