")
")
| Issue |
Med Sci (Paris)
Volume 23, Number 3, Mars 2007
|
|
|---|---|---|
| Page(s) | 303 - 310 | |
| Section | M/S revues | |
| DOI | https://doi.org/10.1051/medsci/2007233303 | |
| Published online | 15 mars 2007 | |
Application des techniques de thérapie génique aux maladies ostéo-articulaires
Gene therapy for osteoarticular disorders
1
Department of orthopaedics and rehabilitation, Gene therapyl aboratory, University of Florida,1600 SW Archer Rd, Rm M2-210,Gainesville, FL 32610, États-Unis
2
Centerfor molecular orthopaedics,Harvard Medical School,221, Longwood Avenue, BLI-152,Boston, MA 02115, États-Unis
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Résumé
L’apparition du concept de gène médicament a considérablement élargi le champ d’application de la thérapie génique. Initialement développées pour les maladies génétiques ou certaines formes de cancer, les stratégies de transfert génique sont désormais étudiées en vue du traitement d’autres pathologies telles que les maladies ostéo-articulaires. La polyarthrite rhumatoïde a ainsi fait l’objet du premier essai clinique de thérapie génique pour le traitement d’une maladie non létale.
Abstract
Osteoarticular disorders are the major cause of disability in Europe and North America. It is estimated that rheumatoid arthritis affects 1 % of the population and that more than two third of people over age 55 develop osteoarthritis. Because there are no satisfactory treatments, gene therapy offers a new therapeutic approach. The delivery of cDNA encoding anti-arthritic proteins to articular cells has shown therapeutic efficacy in numerous animal models in vivo. Through the development and the experimentalprogresses that have been made for both rheumatoid arthritis and osteoarthritis, this review discusses the different gene therapy strategies available today and the safety issues with which they may be associated. Among the different vectors available today, adeno-associated virus seems the best candidate for a direct in vivo gene delivery approach for the treatment of joint disorders.
© 2007 médecine/sciences - Inserm / SRMS
Le développement de traitements par thérapie génique constitue une avancée médicale extraordinaire et suscite beaucoup d’espoir. Deux premiers succès cliniques ont été obtenus en France, puis en Grande-Bretagne, avec la guérison d’enfants atteints d’un déficit immunitaire sévère [1, 2]. Après l’apparition de 3 cas de leucémie dans l’essai français [3], de nombreux progrès ont été faits dans la compréhension des mécanismes responsables de l’apparition d’effets indésirables. Cette connaissance est d’autant plus indispensable lorsque les techniques de transfert génique s’appliquent au traitement de pathologies non létales telles que la plupart des maladies ostéo-articulaires.
Pourquoi avoir développé de nouvelles thérapies pour les maladies ostéo-articulaires ?
Un tiers des populations occidentales serait atteint par les pathologies articulaires inflammatoires et/ou dégénératives, dont les plus fréquentes sont la polyarthrite rhumatoïde (PR) et l’arthrose. Ce type de maladie constitue la première cause d’invalidité chez les personnes âgées de plus de 65 ans. Grâce aux traitements actuels plus puissants et mieux ciblés, le recours à la chirurgie est moins nécessaire et un nombre croissant de patients conserve une activité professionnelle.
L’association entre traitements symptomatiques et traitements de fond reste cependant lourde, et s’accompagne bien souvent d’effets secondaires (Tableau I). Lors d’épisodes aigus, il est nécessaire de calmer la douleur et de juguler l’inflammation. Les prescriptions de corticoïdes et d’anti-inflammatoires non stéroïdiens (AINS) traditionnels sont efficaces lors des périodes inflammatoires sévères, mais ne peuvent pas être prescrits à long terme. Les inhibiteurs spécifiques de la cyclo-oxygénase-2 (« coxibs ») permettent des thérapies plus longues en diminuant les effets indésirables gastro-intestinaux observés avec les AINS traditionnels. Cependant, en février 2005, l’AFSSAPS (Agence française de sécurité sanitaire des produits de santé) a publié une nouvelle liste de contre-indications les concernant : cette information fait suite au retrait du marché du rofécoxib (Vioxx®), intervenant après l’apparition de complications sévères vasculaires, cardiaques ou cérébrales. Dans les pathologies les plus avancées, les traitements de fond tels que les médicaments modifiant l’affection rhumatismale (DMARD, disease modifying anti-rheumatic drugs) et les protéines modificatrices de la réponse biologique (BRM, biologic response modifier) restent essentiels. Les DMARD affaiblissent cependant la réponse du système immunitaire face à une agression extérieure. Quant aux BRM, et notamment aux anti-TNF (tumor necrosis factor), ils ont révolutionné les traitements actuels. Cependant, pour être efficaces, ces traitements nécessitent des injections répétées et fréquentes. L’aspect financier non négligeable, que l’administration s’effectue en ambulatoire ou à l’hôpital, handicape encore leur généralisation : de fait, ce traitement concerne moins d’un quart des patients atteints de PR sévères en France. L’estimation du coût total de ces biothérapies (coûts médicaux directs, coûts non médicaux directs et coûts indirects) oscille en effet entre 15 000 et 20 000 euros par an et par patient, parfois plus.
Traitements actuels disponibles pour les maladies ostéo-articulaires. * Adalimumab, ethanercept et infliximab présentent tous trois des contre-indications (c’est-à-dire tuberculose évolutive, septicémie ou risque de septicémie, insuffisance cardiaque…).
La thérapie génique, en permettant une production constante de la protéine médicament directement dans l’articulation, limiterait de façon considérable le nombre d’injections nécessaires ainsi que le coût global du traitement. De plus, l’activité biologique des protéines transgéniques semble supérieure à celle des protéines recombinantes [4, 5]. In vivo, lors de l’utilisation de l’antagoniste du récepteur de l’interleukine-1 (IL-1) (IL-1Rα) dans un modèle d’arthrite chez le rat, cette différence atteint même un ordre de magnitude 4 [6].
Quels gènes sont les meilleurs candidats ? Pour quels apports thérapeutiques ?
Dans la polyarthrite rhumatoïde
Dans une articulation atteinte de PR, la présence chronique de cytokines inflammatoires telles que l’IL-1 ou le TNFα active les cellules de la membrane synoviale hypertrophiée, induisant la sécrétion de différentes protéines qui érodent l’os et le cartilage environnants. L’élimination des cellules participant au développement de la maladie par surexpression de protéines apoptotiques, notamment, constitue une stratégie thérapeutique intéressante. Ainsi, le transfert intra-articulaire du gène de la thymidine kinase, dont le produit phosphoryle le ganciclovir administré par voie veineuse en un inhibiteur de l’ADN polymérase, induit une cytolyse de la membrane synoviale et une diminution du volume de l’articulation dans un modèle d’arthrite induite par antigène chez le lapin [7], ou par le collagène chez le singe [8]. Un autre exemple de synovectomie génétique a été réalisé en utilisant le transfert de Fas ligand (FasL) dans un modèle d’arthrite au collagène chez la souris [9, 10].
Face à la chronicité des processus impliqués dans la PR, une stratégie bloquant les phases de poussées inflammatoires constitue une autre approche thérapeutique. Chez le rat, l’administration intra-articulaire ou systémique d’un adeno-associated virus de type 2 (AAV-2) exprimant le récepteur du TNFα (p75, TNFR) couplé à une immunoglobuline (TNFR:Fc) (Ethanercept) dans un modèle d’arthrite induite par le streptocoque se traduit par une très forte diminution de l’infiltration cellulaire, de la formation du pannus, de la dégradation des tissus cartilagineux et osseux et de l’expression des ARN messagers de différentes cytokines pro-inflammatoires [11]. Des résultats similaires ont été obtenus après administration intra-articulaire de la forme soluble du TNFR à des souris transgéniques arthritiques exprimant le TNFα [12]. L’injection intra-articulaire de l’ADNc codant pour l’IL-1Rα (Anakinra) via un AAV ou un lentivirus chez le rat réduit l’inflammation de la membrane synoviale et les effets cataboliques sur le cartilage dans des modèles d’arthrites induites respectivement par le LPS [13] ou l’IL-1 [14]. De nombreux autres transgènes (Tableau II) et leurs potentiels anti-arthritiques ont également été testés. Citons notamment ceux codant pour les cytokines anti-inflammatoires IL-4 et IL-10, dont la surexpression prévient la dégradation du cartilage et l’inflammation dans des modèles d’arthrites induites par le collagène chez la souris [15, 16].
Gènes codant pour des protéines potentiellement utiles dans le traitement de la polyarthrite rhumatoïde. * AP-1 bloquant signifie bloqueur du facteur nucléaire AP-1. Ce facteur nucléaire est, entre autre, actif en présence d’inflammation via la voie des MAP-kinases.
Dans l’arthrose
La dégradation du cartilage dans l’arthrose se traduit par une perte des composants matriciels, en particulier des protéoglycanes, associée à un changement qualitatif des fibres de collagène. Des différents médiateurs responsables de la perte des composants matriciels du cartilage, l’IL-1 est de loin le plus important. À faible concentration, l’IL-1 inhibe fortement la synthèse de cartilage, tandis que des concentrations élevées entraînent sa dégradation [17]. La surexpression de gène(s) inhibant les voies signalétiques de l’IL-1 est, de même que pour la PR, attrayante dans le traitement de l’arthrose. L’IL-1Rα seul ou en combinaison avec l’IGF1 (insulin-like growth factor) a donné des résultats intéressants, en bloquant la déplétion en protéoglycanes sur des explants cartilagineux stimulés par l’IL-1 [18]. L’administration de synoviocytes surexprimant l’IL-1Rα dans des modèles d’arthrose par section du ligament croisé antérieur chez le chien s’est traduite par une réduction nette des lésions macroscopiques cartilagineuses au niveau du plateau tibial et des condyles [19]. Des résultats similaires ont été obtenus après injection intra-articulaire directe d’un adénovirus exprimant l’IL-1Rα dans un modèle d’arthrose équine [20]. De manière intéressante, dans un modèle de section du ligament croisé antérieur chez le lapin, l’association avec l’IL-10 augmente le potentiel thérapeutique de l’IL-1Rα [21].
L’utilisation de cellules souches mésenchymateuses (CSM) et de leur potentiel à former des tissus cartilagineux dans différents supports matriciels semble une stratégie prometteuse dans l’optique d’une réparation tissulaire [22, 23]. Différents gènes codant pour des facteurs de croissances (IGF1, BMP [bone morphogenic protein] 2, 4, 6 et 7, TGFβ), des facteurs de transcription (sox-5,-6,-9) ou d’autres molécules (SMAD, Indian Hedgehog, protéines wnt) influencent l’homéostasie du cartilage en favorisant l’anabolisme [24–27]. Des CSM de lapin transduites par un adénovirus transportant l’ADNc codant pour BMP2 ont développé un phénotype chondrocytaire après 3 semaines de culture (matrice extracellulaire riche en collagène de type II et agrécane, absence de collagène de type I) [28]. Ces protéines transgéniques peuvent également être utilisées en combinaison avec des molécules réduisant le catabolisme de la matrice cartilagineuse (inhibiteurs des métalloprotéinases matricielles, notamment). Toutefois, la diffusion systémique de certaines de ces protéines entraîne parfois des effets secondaires plus ou moins graves. Ainsi, la surexpression du TGFβ dans la membrane synoviale provoque l’apparition de fibroses pulmonaires massives nécessitant l’euthanasie de l’animal (données personnelles), tandis que le transfert de l’ADNc exprimant BMP2 a été suivi de formations osseuses ectopiques [29].
Une autre approche utilisant l’accumulation intracellulaire de glucosamine semble protéger les cellules contre les effets néfastes de l’IL-1 [30]. La glucosamine est utilisée de façon empirique dans le traitement de l’arthrose, et bien que ce ne soit pas une protéine, son accumulation est rendue possible par la surexpression de la glutamine fructose 6-phosphate-amido-transférase (GFAT), enzyme impliquée dans sa biosynthèse. In vitro, une inhibition des effets néfastes de l’IL-1 a été observée à la fois sur des synoviocytes et des chondrocytes préalablement transduits par un adénovirus codant pour l’ADNc de la GFAT [31]. In vivo, l’administration intra-articulaire de cet adénovirus prévient la dégradation du cartilage dans un modèle d’arthrite chez le rat (données personnelles).
Traitement des maladies ostéo-articulaires par thérapie génique : quels problèmes, quelles solutions ?
Tout traitement thérapeutique comporte potentiellement un risque pour le patient. La thérapie génique, en ce sens, ne fait pas exception à la règle. Afin de minimiser ces risques, il est indispensable de comprendre les techniques de transfert de gène et les effets biologiques qui y sont associés. La transduction de cellules in vivo est généralement bien tolérée, et les problèmes pathologiques et immunologiques parfois décrits sont majoritairement dus à l’expression de protéines virales. Néanmoins, une réponse immunitaire cellulaire dirigée contre les cellules transduites ou une réponse humorale avec apparition d’anticorps contre la protéine thérapeutique peuvent être observées. La possibilité d’une toxicité de la protéine transgénique n’est pas non plus à exclure. L’utilisation, lorsque cela est possible, d’un transgène dont la pharmacocinétique et la cytotoxicité sont connues (IL-1Ra, sTNFR, etc.) simplifie au moins ce dernier point.
La suspension ou l’adaptation d’un protocole en fonction de l’évolution de la maladie est parfois nécessaire lors d’un traitement thérapeutique. Il est donc important de pouvoir réguler de façon précise et délibérée l’expression du transgène. Le système tetracycline-on (Tet-ON), où la transcription du transgène est dépendante de la présence d’un ligand externe administrable par voie orale, reste l’un des plus étudiés. Son utilisation dans un modèle d’arthrite par le collagène a montré une expression du transgène 10 fois supérieure en présence du ligand (doxycycline) et prévient la progression de la maladie [32]. Les nouvelles versions (Figure 1), utilisant notamment la co-expression de cassettes bi-cistroniques via un IRES (internal ribosome entry site), sont prometteuses. Chez le singe [33] et la souris [34], ces constructions permettent d’obtenir un niveau d’expression basale du transgène négligeable en l’absence de doxycycline, ainsi qu’un contrôle strict de l’expression du transgène sur plusieurs années. Cependant, le potentiel immunogène de ce système fait l’objet de résultats contradictoires [35, 36]. Afin de s’affranchir de ce problème, le système inductible par la rapamycine (Rp) n’utilise que des éléments d’origine humaine [37, 38]. Il est fondé sur la dimérisation de facteurs de transcription en présence de Rp. Bien que tout aussi intéressant que le système Tet-ON en terme d’induction et d’expression basale, ce système nécessite toutefois l’administration de deux vecteurs (Figure 2). De plus, la Rp est immunosuppressive et sa biodisponibilité après ingestion orale limitée [39]. Le développement d’analogues non immunosuppresseurs de la Rp semble encourageant [40]. Enfin, d’autres systèmes présentent également un intérêt pour le traitement par thérapie génique des maladies ostéo-articulaires : citons notamment les promoteurs hybrides, auto-inductibles par un stimulus inflammatoire [41, 42]. Ces constructions sont fondées sur la présence de sites de fixation sur l’ADN pour des facteurs de transcription communs aux différentes voies signalétiques de l’inflammation (NFκB [nuclear factor κB], AP1 [activating protein 1], STAT3 [signal transducer and activator of transcription 3], etc.). Néanmoins, une expression inappropriée du transgène (notamment lors d’une infection) reste possible, et constitue indéniablement un désavantage à l’utilisation de ces systèmes.
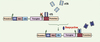 |
Figure 1. Le système inductible par la doxycycline (DOX) [TET-ON (rtTA/tTS)]. En l’absence de DOX, le ligand DOX-dependent trans-silencer (tTS) se fixe sur le promoteur et réprime l’expression du transgène. En présence de DOX, tTS se détache de l’ADN et le transactivateur inversement régulé par le ligand (rtTA) s’y fixe et permet l’expression du transgène. |
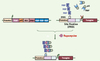 |
Figure 2. Le système inductible par la Rapamycine (Rp). En présence de Rp, il y a formation d’un hétérodimère entre l’immunophiline (FKBP) et la protéine FKBP associée à la Rp (FRAP). Lorsque FKBP est fusionnée avec un zinc-finger homeodomain fusion (ZFDH1) et FRAP à un élément activateur p65, la présence de Rp permet la transcription du transgène sous contrôle d’un promoteur dépendant de la présence de ZFDH1-FKBP. |
Un protocole de thérapie génique efficace se doit d’acheminer sans cytotoxicité la/les protéines transgéniques dans le/les tissus cible(s) à des doses thérapeutiques. De ce fait, les propriétés intrinsèques de chaque vecteur vont orienter, conditionner leurs applications. La faible efficacité de transfection des tissus articulaires in vivo par les vecteurs d’origine non virale [43, 44] oriente désormais un nombre croissant d’équipes scientifiques vers l’utilisation de vecteurs viraux recombinants intégratifs ou non intégratifs (Tableau III). En présence d’une cassette d’expression adéquate, les vecteurs intégratifs [45] permettent le maintien de l’expression de la protéine transgénique au cours des divisions successives de la cellule transduite. Cette intégration peut être intéressante dans le traitement de certaines pathologies chroniques articulaires. En effet, une amplification de l’expression du transgène a été observée lors de la formation du pannus après administration intra-articulaire d’un lentivirus recombinant dans un modèle d’arthrite induite par l’IL-1, suggérant une autorégulation de l’expression transgénique en présence d’inflammation [14]. Le pouvoir infectieux des lentivirus qui, contrairement aux rétrovirus classiques dérivés du Mo-MLV (moloney murine leukemia virus), est indépendant du cycle cellulaire, représente un atout lors de stratégies in vivo en permettant l’injection directe du vecteur dans l’articulation. Cependant, malgré la délétion de toutes les séquences pathogènes et de l’enveloppe du VIH (virus de l’immunodéficience humaine), un obstacle d’ordre psychologique limite encore leur développement clinique pour le traitement de pathologies ostéo-articulaires, la plupart non létales.
Propriétés des différents vecteurs.
Les AAV présentent le profil sanitaire le plus rassurant (Tableau III). Ils ne sont associés à aucune pathologie, sont peu immunogènes et non inflammatoires [46, 47]. Malgré la possibilité d’insertion aléatoire du génome de l’AAV recombinant, cette fréquence reste faible et n’a pas pour l’instant soulevé de questions sanitaires. Il a également été montré qu’une protéine transgénique peut être exprimée plusieurs années après l’administration d’AAV recombinants [48–50], notamment lors de la modification génétique de tissu à faible régénération comme le muscle. Plusieurs points de désaccord persistent néanmoins après administration intra-articulaire d’AAV recombinants, notamment en ce qui concerne l’efficacité de ces vecteurs à transduire une membrane synoviale saine versus enflammée, et quant à la durée d’expression du transgène au sein de l’articulation [51]. Toutefois, un essai clinique de phase I, réalisé chez des patients atteint de PR (Targeted genetics corporation, Seattle, WA, États-Unis), a montré d’une part une bonne tolérance lors de l’administration intra-articulaire de 1011 particules/ml d’un AAV-2 exprimant le TNFR : Fc, et d’autre part une diminution, sur plusieurs mois, du volume des articulations traitées [52]. Le pouvoir de transduction intra-articulaire des AAV recombinants diffère néanmoins d’un sérotype à l’autre en fonction de l’espèce : ainsi, les AAV de type 5 transduisent de manière plus efficace in vitro les synoviocytes humains que les AAV de sérotype 2 [53]. Par ailleurs, le développement de nouveaux variants AAV auto-complémentaires [54] permet l’obtention de niveaux d’expression intra-articulaire élevés, et ce même en absence d’inflammation. Ces concentrations sont de l’ordre du ng/ml de protéine transgénique après injection intra-articulaire de 1011 particules d’AAV-5 ou -8 exprimant le TNFR chez le rat (données personnelles).
Conclusions et perspectives
La connaissance préalable des données cliniques, pharmacocinétiques et de biodistribution des BRM, tels que le Kineret® (IL-1Rα) et l’Enbrel® (sTNFR), a accéléré le développement de protocoles cliniques utilisant le transfert de l’ADNc codant pour ces protéines anti-inflammatoires. La PR a été la première des conditions ostéo-articulaires à être la cible de traitements par thérapie génique. Le premier essai clinique de phase I a été réalisé en 1996 chez 9 femmes dont les stades avancés de PR nécessitaient un remplacement chirurgical des articulations métacarpophalangiennes 2 à 5. Cet essai utilisait une stratégie ex vivo, nécessitant le prélèvement de cellules sur le patient, leur modification génétique en laboratoire par un rétrovirus exprimant l’IL-1Rα, puis leur réimplantation dans les articulations atteintes. Une semaine après, l’expression du transgène a été détectée et aucun effet secondaire n’a été observé [55]. Dix ans se sont écoulés depuis, et 8 essais cliniques impliquant des techniques de transfert de gène ont été réalisés [51]. Cinq des 8 essais (tous de phase I, dont celui de la Targeted genetics corporation) étaient fondés sur l’administration locale intra-articulaire de vecteur. Dans les 3 autres essais (phases I, IIa ou II), il s’agissait d’une administration intraveineuse ou sous-cutanée d’ARN antisens ciblant le TNF. Bien que les résultats finaux n’aient pas encore été publiés, les critères sanitaires ont dans tous les cas été remplis et aucun effet secondaire n’a pour le moment été reporté. Tous ces essais cliniques ciblaient cependant la PR et, en dépit du nombre croissant de personnes atteintes d’arthrose et de l’absence de « médicaments » efficaces, aucune étude clinique de thérapie génique appliquée au traitement de cette maladie n’a pour l’instant était conduite. Toutefois, un essai de phase II fondé sur l’injection intra-articulaire d’un AAV exprimant l’IL-1Rα dans une articulation arthrosique est actuellement en préparation (communication personnelle du Pr Evans).
Les résultats prometteurs obtenus notamment pour le traitement de la PR ont permis l’application de techniques de transfert de gène à d’autres pathologies du système locomoteur (Tableau IV). Au vu des résultats encourageants, les essais cliniques par transfert de gène(s) pour les différentes pathologies ostéo-articulaires devraient se multiplier dans un proche avenir.
Autres applications ostéo-articulaires possibles de la thérapie génique.

Article reçu le 6 février 2006, accepté le 15 juillet 2006.
Références
- Cavazzana-Calvo M, Hacein-Bey S, de Saint Basile G, et al. Gene therapy of human severe combined immunodeficiency (SCID)-X1 disease. Science 2000; 288 : 669–72. [Google Scholar]
- Gaspar HB, Parsley KL, Howe S, et al. Gene therapy of X-linked severe combined immunodeficiency by use of a pseudotyped gammaretroviral vector. Lancet 2004; 364 : 2181–7. [Google Scholar]
- Hacein-Bey-Abina S, Von Kalle C, Schmidt M, et al. LMO2-associated clonal T cell proliferation in two patients after gene therapy for SCID-X1. Science 2003; 302 : 415–9. [Google Scholar]
- Palmer GD, Steinert A, Pascher A, et al. Gene-induced chondrogenesis of primary mesenchymal stem cells in vitro. Mol Ther 2005; 12 : 219–28. [Google Scholar]
- Gouze JN, Gouze E, Palmer GD, et al. A comparative study of the inhibitory effects of interleukin-1 receptor antagonist following administration as a recombinant protein or by gene transfer. Arthritis Res Ther 2003; 5 : R301–9. [Google Scholar]
- Makarov SS, Olsen JC, Johnston WN, et al. Suppression of experimental arthritis by gene transfer of interleukin 1 receptor antagonist cDNA. Proc Natl Acad Sci USA 1996; 93 : 402–6. [Google Scholar]
- Sant SM, Suarez TM, Moalli MR, et al. Molecular lysis of synovial lining cells by in vivo herpes simplex virus-thymidine kinase gene transfer. Hum Gene Ther 1998; 9 : 2735–43. [Google Scholar]
- Goossens PH, Schouten GJ, ’t Hart BA, et al. Feasibility of adenovirus-mediated nonsurgical synovectomy in collagen-induced arthritis-affected rhesus monkeys. Hum Gene Ther 1999; 10 : 1139–49. [Google Scholar]
- Yao Q, Glorioso JC, Evans CH, et al. Adenoviral mediated delivery of FAS ligand to arthritic joints causes extensive apoptosis in the synovial lining. J Gene Med 2000; 2 : 210–9. [Google Scholar]
- Zhang H, Yang Y, Horton JL, et al. Amelioration of collagen-induced arthritis by CD95 (Apo-1/Fas)-ligand gene transfer. J Clin Invest 1997; 100 : 1951–7. [Google Scholar]
- Chan JM, Villarreal G, Jin WW, et al. Intraarticular gene transfer of TNFR : Fc suppresses experimental arthritis with reduced systemic distribution of the gene product. Mol Ther 2002; 6 : 727–36. [Google Scholar]
- Zhang HG, Xie J, Yang P, et al. Adeno-associated virus production of soluble tumor necrosis factor receptor neutralizes tumor necrosis factor alpha and reduces arthritis. Hum Gene Ther 2000; 11 : 2431–42. [Google Scholar]
- Pan RY, Chen SL, Xiao X, et al. Therapy and prevention of arthritis by recombinant adeno-associated virus vector with delivery of interleukin-1 receptor antagonist. Arthritis Rheum 2000; 43 : 289–97. [Google Scholar]
- Gouze E, Pawliuk R, Gouze JN, et al. Lentiviral-mediated gene delivery to synovium : potent intra-articular expression with amplification by inflammation. Mol Ther 2003; 7 : 460–6. [Google Scholar]
- Lubberts E, Joosten LA, van Den Bersselaar L, et al. Adenoviral vector-mediated overexpression of IL-4 in the knee joint of mice with collagen-induced arthritis prevents cartilage destruction. J Immunol 1999; 163 : 4546–56. [Google Scholar]
- Setoguchi K, Misaki Y, Araki Y, et al. Antigen-specific T cells transduced with IL-10 ameliorate experimentally induced arthritis without impairing the systemic immune response to the antigen. J Immunol 2000; 165 : 5980–6. [Google Scholar]
- Goldring MB, Birkhead JR, Suen LF, et al. Interleukin-1 beta-modulated gene expression in immortalized human chondrocytes. J Clin Invest 1994; 94 : 2307–16. [Google Scholar]
- Nixon AJ, Haupt JL, Frisbie DD, et al. Gene-mediated restoration of cartilage matrix by combination insulin-like growth factor-I/interleukin-1 receptor antagonist therapy. Gene Ther 2005; 12 : 177–86. [Google Scholar]
- Pelletier JP, Caron JP, Evans C, et al. In vivo suppression of early experimental osteoarthritis by interleukin-1 receptor antagonist using gene therapy. Arthritis Rheum 1997; 40 : 1012–9. [Google Scholar]
- Frisbie DD, Ghivizzani SC, Robbins PD, et al. Treatment of experimental equine osteoarthritis by in vivo delivery of the equine interleukin-1 receptor antagonist gene. Gene Ther 2002; 9 : 12–20. [Google Scholar]
- Zhang X, Mao Z, Yu C. Suppression of early experimental osteoarthritis by gene transfer of interleukin-1 receptor antagonist and interleukin-10. J Orthop Res 2004; 22 : 742–50. [Google Scholar]
- Li WJ, Tuli R, Okafor C, et al. A three-dimensional nanofibrous scaffold for cartilage tissue engineering using human mesenchymal stem cells. Biomaterials 2005; 26 : 599–609. [Google Scholar]
- Pascher A, Palmer GD, Steinert A, et al. Gene delivery to cartilage defects using coagulated bone marrow aspirate. Gene Ther 2004; 11 : 133–41. [Google Scholar]
- Nakamura Y, Nawata M, Wakitani S. Expression profiles and functional analyses of Wnt-related genes in human joint disorders. Am J Pathol 2005; 167 : 97–105. [Google Scholar]
- Tyler JA. Insulin-like growth factor 1 can decrease degradation and promote synthesis of proteoglycan in cartilage exposed to cytokines. Biochem J 1989; 260 : 543–8. [Google Scholar]
- Lafeber FP, van Roy HL, van der Kraan PM, et al. Transforming growth factor-beta predominantly stimulates phenotypically changed chondrocytes in osteoarthritic human cartilage. J Rheumatol 1997; 24 : 536–42. [Google Scholar]
- Tchetina EV, Squires G, Poole AR. Increased type II collagen degradation and very early focal cartilage degeneration is associated with upregulation of chondrocyte differentiation related genes in early human articular cartilage lesions. J Rheumatol 2005; 32 : 876–86. [Google Scholar]
- Palmer G, Pascher A, Gouze E, et al. Development of gene-based therapies for cartilage repair. Crit Rev Eukaryot Gene Expr 2002; 12 : 259–73. [Google Scholar]
- Gelse K, Jiang QJ, Aigner T, et al. Fibroblast-mediated delivery of growth factor complementary DNA into mouse joints induces chondrogenesis but avoids the disadvantages of direct viral gene transfer. Arthritis Rheum 2001; 44 : 1943–53. [Google Scholar]
- Gouze JN, Bianchi A, Becuwe P, et al. Glucosamine modulates IL-1-induced activation of rat chondrocytes at a receptor level, and by inhibiting the NF-kappa B pathway. FEBS Lett 2002; 510 : 166–70. [Google Scholar]
- Gouze JN, Gouze E, Palmer GD, et al. Adenovirus-mediated gene transfer of glutamine : fructose-6-phosphate amidotransferase antagonizes the effects of interleukin-1beta on rat chondrocytes. Osteoarthritis Cartilage 2004; 12 : 217–24. [Google Scholar]
- Perez N, Plence P, Millet V, et al. Tetracycline transcriptional silencer tightly controls transgene expression after in vivo intramuscular electrotransfer : application to interleukin 10 therapy in experimental arthritis. Hum Gene Ther 2002; 13 : 2161–72. [Google Scholar]
- Lamartina S, Silvi L, Roscilli G, et al. Construction of an rtTA2(s)-m2/tts(kid)-based transcription regulatory switch that displays no basal activity, good inducibility, and high responsiveness to doxycycline in mice and non-human primates. Mol Ther 2003; 7 : 271–80. [Google Scholar]
- Salucci V, Scarito A, Aurisicchio L, et al. Tight control of gene expression by a helper-dependent adenovirus vector carrying the rtTA2(s)-M2 tetracycline transactivator and repressor system. Gene Ther 2002; 9 : 1415–21. [Google Scholar]
- Favre D, Blouin V, Provost N, et al. Lack of an immune response against the tetracycline-dependent transactivator correlates with long-term doxycycline-regulated transgene expression in nonhuman primates after intramuscular injection of recombinant adeno-associated virus. J Virol 2002; 76 : 11605–11. [Google Scholar]
- Latta-Mahieu M, Rolland M, Caillet C, et al. Gene transfer of a chimeric trans-activator is immunogenic and results in short-lived transgene expression. Hum Gene Ther 2002; 13 : 1611–20. [Google Scholar]
- Wang J, Voutetakis A, Papa M, et al. Rapamycin control of transgene expression from a single AAV vector in mouse salivary glands. Gene Ther 2006; 13 : 187–90. [Google Scholar]
- Rivera VM, Clackson T, Natesan S, et al. A humanized system for pharmacologic control of gene expression. Nat Med 1996; 2 : 1028–32. [Google Scholar]
- Xu ZL, Mizuguchi H, Mayumi T, Hayakawa T. Regulated gene expression from adenovirus vectors : a systematic comparison of various inducible systems. Gene 2003; 309 : 145–51. [Google Scholar]
- Chong H, Ruchatz A, Clackson T, et al. A system for small-molecule control of conditionally replication-competent adenoviral vectors. Mol Ther 2002; 5 : 195–203. [Google Scholar]
- van de Loo FA, de Hooge AS, Smeets RL, et al. An inflammation-inducible adenoviral expression system for local treatment of the arthritic joint. Gene Ther 2004; 11 : 581–90. [Google Scholar]
- Bakker AC, van de Loo FA, Joosten LA, et al. C3-Tat/HIV-regulated intraarticular human interleukin-1 receptor antagonist gene therapy results in efficient inhibition of collagen-induced arthritis superior to cytomegalovirus-regulated expression of the same transgene. Arthritis Rheum 2002; 46 : 1661–70. [Google Scholar]
- Li S, Ma Z. Nonviral gene therapy. Curr Gene Ther 2001; 1 : 201–26. [Google Scholar]
- Evans CH, Ghivizzani SC, Robbins PD. The 2003 Nicolas Andry Award. Orthopaedic gene therapy. Clin Orthop Relat Res 2004; 316–29. [Google Scholar]
- Kim SH, Kim S, Robbins PD. Retroviral vectors. Adv Virus Res 2000; 55 : 545–63. [Google Scholar]
- Smith-Arica JR, Bartlett JS. Gene therapy : recombinant adeno-associated virus vectors. Curr Cardiol Rep 2001; 3 : 43–9. [Google Scholar]
- Zhang YC, Powers M, Wasserfall C, et al. Immunity to adeno-associated virus serotype 2 delivered transgenes imparted by genetic predisposition to autoimmunity. Gene Ther 2004; 11 : 233–40. [Google Scholar]
- Bouchard S, MacKenzie TC, Radu AP, et al. Long-term transgene expression in cardiac and skeletal muscle following fetal administration of adenoviral or adeno-associated viral vectors in mice. J Gene Med 2003; 5 : 941–50. [Google Scholar]
- Rivera VM, Gao GP, Grant RL, et al. Long-term pharmacologically regulated expression of erythropoietin in primates following AAV-mediated gene transfer. Blood 2005; 105 : 1424–30. [Google Scholar]
- Stieger K, Le Meur G, Lasne F, et al. Long-term doxycycline-regulated transgene expression in the retina of nonhuman primates following subretinal injection of recombinant AAV vectors. Mol Ther 2006; 13 : 967–75. [Google Scholar]
- Evans CH, Ghivizzani SC, Robbins PD. Gene therapy for arthritis : What next ? Arthritis Rheum 2006; 54 : 1714–29. [Google Scholar]
- Heald A, Pate G. Clinical studies of intra-articular administration of a recombinant adeno-associated vector containing a TNF-antagonist gene in inflammatory arthritis. Baltimore : ASGT 9th Annual Meeting, 2006. [Google Scholar]
- Apparailly F, Khoury M, Vervoordeldonk MJ, et al. Adeno-associated virus pseudotype 5 vector improves gene transfer in arthritic joints. Hum Gene Ther 2005; 16 : 426–34. [Google Scholar]
- McCarty DM, Fu H, Monahan PE, et al. Adeno-associated virus terminal repeat (TR) mutant generates self-complementary vectors to overcome the rate-limiting step to transduction in vivo. Gene Ther 2003; 10 : 2112–8. [Google Scholar]
- Evans CH, Robbins PD, Ghivizzani SC, et al. Gene transfer to human joints : progress toward a gene therapy of arthritis. Proc Natl Acad Sci USA 2005; 102 : 8698–703. [Google Scholar]
Liste des tableaux
Traitements actuels disponibles pour les maladies ostéo-articulaires. * Adalimumab, ethanercept et infliximab présentent tous trois des contre-indications (c’est-à-dire tuberculose évolutive, septicémie ou risque de septicémie, insuffisance cardiaque…).
Gènes codant pour des protéines potentiellement utiles dans le traitement de la polyarthrite rhumatoïde. * AP-1 bloquant signifie bloqueur du facteur nucléaire AP-1. Ce facteur nucléaire est, entre autre, actif en présence d’inflammation via la voie des MAP-kinases.
Liste des figures
|
Figure 1. Le système inductible par la doxycycline (DOX) [TET-ON (rtTA/tTS)]. En l’absence de DOX, le ligand DOX-dependent trans-silencer (tTS) se fixe sur le promoteur et réprime l’expression du transgène. En présence de DOX, tTS se détache de l’ADN et le transactivateur inversement régulé par le ligand (rtTA) s’y fixe et permet l’expression du transgène. |
| Dans le texte | |
|
Figure 2. Le système inductible par la Rapamycine (Rp). En présence de Rp, il y a formation d’un hétérodimère entre l’immunophiline (FKBP) et la protéine FKBP associée à la Rp (FRAP). Lorsque FKBP est fusionnée avec un zinc-finger homeodomain fusion (ZFDH1) et FRAP à un élément activateur p65, la présence de Rp permet la transcription du transgène sous contrôle d’un promoteur dépendant de la présence de ZFDH1-FKBP. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.