")
")
| Issue |
Med Sci (Paris)
Volume 42, Number 5, Mai 2026
|
|
|---|---|---|
| Page(s) | 427 - 431 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/2026083 | |
| Published online | 29 mai 2026 | |
HEVidence à la porte
Le mystère de l’entrée du virus de l’hépatite E dans les cellules se dévoile
Knocking on the cell’s door: HEVidence of how HEV gets in
Université de Lille, CNRS, Inserm, CHU de Lille, Institut Pasteur de Lille, U1019-UMR 9017, Centre d’infection et d’immunité de Lille (CIIL), Lille, France
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Le virus de l’hépatite E (HEV) constitue aujourd’hui la principale cause d’hépatite virale aigüe dans le monde, avec environ 20 millions d’infections et 40 000 à 70 000 décès par an. Ce virus demeure largement sous-estimé, alors qu’il figure parmi les six principales menaces zoonotiques en termes de risque de propagation dans la population humaine [1]. Bien que l’infection par HEV soit généralement spontanément résolutive, elle peut évoluer vers une forme chronique chez les personnes immunodéprimées. De plus, chez les femmes enceintes ou les personnes atteintes d’une maladie hépatique préexistante, le taux de mortalité associée à cette infection est particulièrement élevé [2]. Un vaccin existe, mais il n’est actuellement utilisé qu’en Chine et au Pakistan. À ce jour, aucun traitement antiviral spécifique n’est disponible, une situation qui s’explique notamment par les nombreuses lacunes dans la compréhension du cycle infectieux du virus. Il est donc nécessaire d’approfondir nos connaissances afin d’identifier de nouvelles cibles thérapeutiques contre l’infection [2].
HEV est un virus à ARN simple brin de polarité positive, qui circule sous deux formes : une forme non enveloppée (nHEV), excrétée dans les selles des personnes infectées et responsable de la propagation du virus dans la population, et une forme « quasi-enveloppée » (eHEV), libérée dans le sang des personnes infectées et probablement responsable de la propagation du virus au sein de l’hôte. Ces deux formes pénètrent dans les cellules par des mécanismes distincts (Figure 1). Plusieurs facteurs cellulaires favorisant la liaison et l’internalisation de nHEV, de eHEV ou de ces deux formes du virus ont été décrits [3]. La liaison initiale de nHEV à la membrane plasmique implique notamment les protéoglycanes à héparane sulfate (HSPG) ainsi que le récepteur des asialoglycoprotéines (ASGPR). Pour eHEV, le récepteur de la phosphatidylsérine TIM1 (T-cell immunoglobulin and mucin domain protein 1) est le seul facteur d’entrée dans la cellule connu à ce jour [4]. Les particules virales, enveloppées ou non, sont ensuite internalisées par une voie d’endocytose dépendante de la clathrine. Plusieurs facteurs de la cellule hôte, tels que le récepteur du facteur de croissance épidermique (EGFR), la tyrosine kinase YES1, et la protéine de choc thermique 90 (HSP90), participent au transport intracellulaire des virions [5-7]. Cependant, des résultats contradictoires ont été publiés concernant le trafic endosomique de HEV. Yin et al. ont proposé que eHEV transite par des compartiments endosomiques Rab5+ et Rab7+, tandis que le transport de nHEV n’utiliserait pas la voie des endosomes [3]. Au contraire, Holla et al. ont montré que nHEV emprunte la voie endosomique Rab5+ et atteint les lysosomes [8].
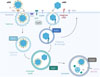 |
Figure 1 Voies d’entrée des particules eHEV et nHEV dans les hépatocytes. Les particules eHEV et nHEV pénètrent dans les hépatocytes par des voies différentes. La phosphatidylsérine (PS) présente dans l’enveloppe virale de eHEV interagit avec le récepteur TIM1 pour permettre l’entrée du virus. Pour nHEV, différentes protéines de l’hôte, notamment les protéoglycanes à héparane sulfate (HSPG), le récepteur de l’asialoglycoprotéine (ASGPR), et les intégrines, peuvent faciliter son entrée. De plus, le récepteur du facteur de croissance épidermique (EGFR) et la tyrosine kinase YES1 jouent également un rôle dans les étapes d’entrée des deux types de particules. Fu et al. ont montré que eHEV transite probablement par les endosomes précoces (Rab5+) et tardifs (Rab7+) pour atteindre les lysosomes (Lamp1+), tandis que nHEV transite préférentiellement par les endosomes de recyclage (Rab11+) et les endosomes tardifs avant d’atteindre les lysosomes. Dans ce dernier compartiment, la membrane virale de eHEV est dégradée par des lipases et la capside par les cathepsines (Pac-mans verts), puis l’ARN viral est libéré dans le cytoplasme, ce qui permet la transcription et la traduction du génome viral. Figure réalisée avec BioRender. |
Récemment, l’équipe dirigée par Viet Loan Dao Thi a utilisé une approche d’imagerie fondée sur la technique RNAscope, une méthode d’hybridation in situ par fluorescence de l’ARN, pour mettre en évidence l’implication de l’intégrine β1 (ITGB1) dans l’internalisation et le transport endocytaire de nHEV [9].
Parmi les facteurs d’entrée de HEV décrits précédemment figure également l’intégrine α3 (ITGA3) [10]. Les chercheurs ont réalisé des analyses protéomiques sur des cellules permissives à l’infection par HEV et ont montré que les cellules d’hépatome PLC/PRF/5, utilisées dans l’étude initiale de Shiota et al. [10], expriment effectivement ITGA3, mais que d’autres cellules d’hépatome (HepG2, Huh-1, Huh-7) fréquemment utilisées et plus permissives à l’infection par le virus, ne l’expriment pas ou très peu. En revanche, elles expriment fortement ITGB1, majoritairement localisée à la membrane plasmique [9]. Puisque que ITGB1 peut former des hétérodimères fonctionnels avec ITGA3, les chercheurs ont voulu préciser le rôle de ITGB1 dans le cycle de HEV.
Ils ont d’abord montré que l’infection par nHEV était réduite dans des cellules S10-3 privées de ITGB1 par invalidation génique, tandis que l’infection par eHEV demeurait inchangée. Ils ont ensuite confirmé ce résultat en utilisant d’autres lignées cellulaires (Huh-7, HepG2/C3A, A549 et Caco-2) transfectées avec un ARN interférent (short interfering RNA, siRNA) ciblant ITGB1. D’autres expériences ont par ailleurs permis de montrer que ITGB1 n’intervient pas dans la réplication du virus. Parmi les différentes fonctions des intégrines figure leur rôle de récepteur du collagène, qui repose sur la reconnaissance d’un motif RGD (arginine, glycine, acide aspartique) présent dans leurs ligands. Dans une expérience fondée sur la compétition, l’ajout d’un peptide RGD aux cellules avant l’infection a entraîné une réduction dose-dépendante de l’infection par nHEV, et ce dans plusieurs lignées cellulaires [9]. Les résultats de ces premières expériences étaient donc en faveur d’un rôle clé de ITGB1 dans l’entrée de nHEV, mais pas dans celle de eHEV.
L’analyse de l’étape d’entrée de HEV dans les cellules a longtemps été limitée par l’absence de méthodes fiables. Les chercheurs ont donc développé une approche d’imagerie innovante fondée sur la technique RNAscope, qui permet d’étudier les particules virales entrantes en détectant les ARN viraux individuels à l’aide de sondes spécifiques (Figure 2). Pour cela, un marquage séquentiel est réalisé par hybridation de ces sondes à leur ARN cible, dans le cas présent l’ARN de HEV, suivie de l’ajout d’amplificateurs et d’autres sondes fluorescentes. Grâce à cette méthode, qui permet d’amplifier le signal spécifique tout en réduisant le bruit de fond, il devient possible de détecter des molécules individuelles. Les chercheurs ont utilisé deux sondes, l’une ciblant ORF1 et l’autre ciblant ORF2, afin d’assurer la détection de particules virales complètes plutôt que de génomes tronqués ou défectueux. Grâce au marquage simultané de ITGB1 à la membrane plasmique par immunofluorescence, les génomes de nHEV et eHEV ont pu être détectés soit à la périphérie, soit à l’intérieur des cellules. Une quantification du « nombre de particules HEV par cellule » a été réalisée en calculant le rapport entre le nombre de points fluorescents correspondant aux génomes HEV et le nombre de noyaux cellulaires par image analysée [9].
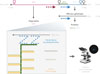 |
Figure 2 Technique d’imagerie RNAscope pour l’ARN de HEV. Le génome du virus de l’hépatite E (HEV) est constitué d’un ARN de 7,2 kilobases (kb) organisé en trois cadres ouverts de lecture (ORF) codant trois protéines virales. ORF1 code une polyprotéine impliquée dans la réplication virale, ORF2 code la protéine de capside, et ORF3 code une petite phosphoprotéine impliquée dans la sécrétion des particules virales. Les protéines ORF2 et ORF3 sont produites à partir d’un ARN sous-génomique de 2,2 kb. RNC : région non codante. Une représentation schématique de la technique RNAscope est présentée. Cette technique d’hybridation in situ « multiplexée » permet la détection spécifique de l’ARN par hybridation de paires de sondes dites « ZZ » sur des séquences cibles adjacentes. Fu et al. ont utilisé deux sondes différentes : une sonde ciblant une région conservée de ORF1, permettant de détecter spécifiquement l’ARN génomique de HEV (flèche en trait pointillé rouge bordeaux ), et une sonde ciblant une région chevauchante de ORF2 et ORF3, permettant de détecter à la fois l’ARN génomique et l’ARN sous-génomique de HEV (flèche en trait pointillé bleu). La combinaison de ces deux sondes permet de distinguer les génomes complets de HEV, encapsidés dans les particules virales, des génomes défectueux, tronqués ou sous-génomiques. Cette distinction est importante pour étudier le processus d’entrée du virus dans la cellule hôte. Après hybridation des paires ZZ, un système d’amplification du signal est assuré par la liaison séquentielle de molécules pré-amplificatrices et amplificatrices, suivie de l’hybridation de multiples sondes fluorescentes, produisant un signal ponctué, spécifique et sensible, correspondant à des molécules individuelles d’ARN. Figure réalisée avec BioRender. |
Le test d’entrée du virus dans la cellule comprenait une étape à la température de 4 °C, permettant uniquement l’attachement des particules virales à la surface cellulaire, suivie soit d’une fixation, soit d’une exposition des cellules à la température de 37 °C afin d’induire l’internalisation. Les chercheurs ont montré que le nombre de particules nHEV liées à la membrane plasmique et internalisées était réduit dans les cellules génétiquement dépourvues de ITGB1 ou dans celles transfectées avec un siRNA ciblant ITGB1. De plus, la réexpression de ITGB1 a permis de rétablir l’entrée du virus. L’extinction, par la technique siRNA, de la kinase d’adhérence focale (FAK), une protéine activée par les intégrines β, a également entraîné une réduction de l’internalisation de nHEV [9].
La possibilité d’une interaction entre la protéine de capside ORF2 et ITGB1 a ensuite été explorée. Les chercheurs ont d’abord montré la colocalisation des particules nHEV et de ITGB1 à la membrane cellulaire. Un motif RGD inversé (DGR) a été identifié dans le domaine saillant de la protéine de capside ORF2. Bien que ce type de motif présente en principe une faible affinité pour les intégrines, l’acide aspartique (D) du motif DGR peut subir une désamination non enzymatique en acide isoaspartique, transformant alors ce motif en un peptide compétitif capable d’interagir avec les sites de liaison des intégrines. La mutation de l’acide aspartique (D) du motif DGR de ORF2 en acide glutamique (E) a entraîné une réduction de l’infectiosité du virus, avec une déficience de la liaison à la cellule et de l’internalisation, confirmant ainsi l’implication de ce motif dans l’entrée de nHEV et son interaction avec ITGB1. Des modélisations par AlphaFold2 ainsi que des tests de ligation de proximité ont apporté des éléments supplémentaires en faveur de l’importance du motif DGR de la protéine de capside dans l’interaction du virus avec ITGB1 [9].
Dans une autre série d’expériences, la cinétique de décapsidation virale a été analysée en combinant la technique RNAscope pour la détection du génome viral et le marquage de la capside par des anticorps. Dans ces conditions, la colocalisation du génome viral et de la capside correspond à des particules infectieuses intactes, tandis que la détection du génome viral seul indique que la décapsidation a eu lieu. Pour les particules nHEV, aucune décapsidation n’a été constatée aux temps précoces de l’infection, mais 24 heures plus tard, l’ensemble des particules nHEV étaient décapsidées. Pour les particules eHEV, la décapsidation se produisait plus lentement [9].
Il avait été précédemment suggéré que HEV est internalisé par une voie d’endocytose dépendante de la clathrine [8]. Les chercheurs ont émis l’hypothèse que nHEV emprunte cette voie en interagissant avec ITGB1, qui est dégradée ou recyclée après l’interaction avec son ligand. Le traitement des cellules avec des inhibiteurs d’acidification des endosomes, tels que la bafilomycine A1 et la concanamycine A, a entraîné une accumulation des particules virales intactes de nHEV et eHEV, indiquant qu’aucune décapsidation ne se produit en l’absence d’acidification des endosomes [9].
D’autres voies de trafic intracellulaire ont ensuite été explorées par inhibition des marqueurs des endosomes précoces, tardifs, et de recyclage (Rab5a, Rab7, et Rab11, respectivement). Des expériences de surexpression de ces protéines combinée au marquage des particules nHEV ou eHEV ont également été réalisées. Les chercheurs ont ainsi montré que nHEV transite préférentiellement par les endosomes de recyclage (Rab11+) et les endosomes tardifs (Rab7+). De plus, nHEV est colocalisé avec ITGB1 dans les compartiments Rab11+, ce qui étaye l’hypothèse selon laquelle l’interaction avec ITGB1 dirige les particules nHEV vers cette voie de trafic intracellulaire. Au contraire, eHEV transite principalement par les endosomes précoces (Rab5+) et les endosomes tardifs (Rab7+) [9] (Figure 1).
Le transport des particules nHEV et eHEV vers les lysosomes, principale destination de la cargaison endocytaire, a ensuite été étudié. La colocalisation du marqueur LAMP1 des lysosomes et de l’ARN viral a permis d’établir que nHEV et eHEV atteignent les lysosomes environ 7 heures et 10 heures après leur internalisation, respectivement. Enfin, des expériences de traitement avec l’inhibiteur de cathepsines E64 ont montré que les cathepsines des lysosomes sont nécessaires à la libération du génome viral, tant pour nHEV que pour eHEV [9]. L’ensemble des résultats expérimentaux obtenus par ces chercheurs les conduisent à suggérer que les intégrines α ne constituent pas un facteur déterminant pour l’entrée de nHEV dans les cellules, mais que celle-ci dépend principalement de ITGB1, une intégrine ubiquitaire et plus abondante à la membrane plasmique. Il est probable que, dans les hépatocytes, ITGB1 forme des hétérodimères avec certaines intégrines α dont l’expression est spécifique de tissu. Il conviendra donc de déterminer si ITGB1 s’associe à des intégrines α spécifiques, dont la présence pourrait jouer un rôle critique dans l’infection par HEV d’autres organes que le foie, tels que le cerveau et l’intestin [11].
Les résultats obtenus par ces chercheurs ont également permis de mieux caractériser les voies d’endocytose empruntées par les particules nHEV et eHEV. L’interaction entre la protéine de capside et ITGB1 induit l’internalisation des particules nHEV et leur adressage vers les endosomes de recyclage, probablement via le recrutement de la protéine FAK. En revanche, les particules eHEV empruntent la voie d’endocytose classique, en transitant par les endosomes précoces. Les particules nHEV ont ensuite été détectées dans des compartiments LAMP1+, correspondant probablement à des endolysosomes. Notons qu’un trafic entre les endosomes de recyclage et les endolysosomes a déjà été décrit pour un certain nombre de récepteurs et de virus, et pourrait être associé à la formation d’autophagosomes [12]. Il reste à déterminer si ce mécanisme contribue également à la sortie de HEV des compartiments endosomiques. Finalement, eHEV et nHEV atteignent tous deux les lysosomes, où les cathepsines clivent la capside virale et exposent des peptides susceptibles de favoriser la fusion des membranes. Ce processus permet la libération du génome viral dans le cytosol, où il devient accessible pour les étapes ultérieures de transcription et de traduction [13].
Il avait été montré précédemment que les protéines Rab5 et Rab11 sont impliquées dans la réplication et l’assemblage de HEV [14, 15], ce qui souligne l’étroite interconnexion des différentes étapes du cycle viral puisqu’on sait désormais que ces protéines sont également impliquées dans le trafic des particules virales après leur entrée dans la cellule. Ce constat justifie la nécessité de disposer de méthodes permettant de distinguer les différentes étapes du cycle viral. La stratégie de co-marquage du génome viral et de la protéine de capside, utilisée par les chercheurs sur une période de 8 à 24 heures correspondant aux étapes précoces de l’infection, répond à cet objectif en permettant la distinction entre les particules virales encore intactes et les génomes décapsidés.
De nombreuses questions subsistent encore quant aux mécanismes moléculaires de l’entrée des particules eHEV et nHEV dans la cellule hôte, ainsi que les raisons pour lesquelles ces étapes précoces sont particulièrement lentes pour HEV comparativement à d’autres virus. Les avancées de la recherche sur ces questions devraient ouvrir la voie à de nouvelles stratégies thérapeutiques contre l’infection par ce virus.
Liens d’intérêt
Les auteurs déclarent ne pas avoir de lien d’intérêt.
Références
- Grange ZL, Goldstein T, Johnson CK, et al. Ranking the risk of animal-to-human spillover for newly discovered viruses. Proc Natl Acad Sci USA 2021 ; 118 : e2002324118. [Google Scholar]
- Corneillie L, Mézière L, Montpellier C, et al. Update on the molecular and cellular biology of hepatitis E virus and therapeutic opportunities. Antivir Res 2026 ; 106353. [Google Scholar]
- Yin X, Ambardekar C, Lu Y, et al. Distinct entry mechanisms for non-enveloped and quasi-enveloped hepatitis E virus. J Virol 2016 ; JVI.02804-15. [Google Scholar]
- Corneillie L, Lemmens I, Montpellier C, et al. The phosphatidylserine receptor TIM1 promotes infection of enveloped hepatitis E virus. Cell Mol Life Sci 2023 ; 80 : 326. [Google Scholar]
- Zheng ZZ, Miao J, Zhao M, et al. Role of heat-shock protein 90 in hepatitis E virus capsid trafficking. J Gen Virol 2010 ; 91 : 1728-36. [Google Scholar]
- Schrader JA, Burkard TL, Brüggemann Y, et al. EGF receptor modulates HEV entry in human hepatocytes. Hepatology 2023 ; 77 : 2104-17. [Google Scholar]
- Haase JA, Baheerathan A, Zhang X, et al. The tyrosine kinase Yes1 is a druggable host factor of HEV. Hepatol Commun 2024 ; 8 : e0553. [Google Scholar]
- Holla P, Ahmad I, Ahmed Z, et al. Hepatitis E virus enters liver cells through a dynamin-2, clathrin and membrane cholesterol-dependent pathway. Traffic 2015 ; 16 : 398-416. [Google Scholar]
- Fu R, Jordan P, Engels Z, et al. Integrin b1 facilitates non-enveloped hepatitis E virus cell entry through the recycling endosome. Nat Commun 2025 ; 16 : 5403. [Google Scholar]
- Shiota T, Li TC, Nishimura Y, et al. Integrin α3 is involved in non-enveloped hepatitis E virus infection. Virology 2019 ; 536 : 119-24. [Google Scholar]
- Kamar N, Marion O, Abravanel F, et al. Extrahepatic manifestations of hepatitis E virus. Liver Int 2016 ; 36 : 467-72. [CrossRef] [PubMed] [Google Scholar]
- Xu J, Gu J, Pei W, et al. The role of lysosomal membrane proteins in autophagy and related diseases. FEBS J 2024 ; 291 : 3762-85. [Google Scholar]
- Klöhn M, Burkard T, Janzen J, et al. Targeting cellular cathepsins inhibits hepatitis E virus entry. Hepatology 2024 ; 80 : 1239-51. [Google Scholar]
- Bentaleb C, Hervouet K, Montpellier C, et al. The endocytic recycling compartment serves as a viral factory for hepatitis E virus. Cell Mol Life Sci 2022 ; 79 : 615. [Google Scholar]
- Oechslin N, Silva ND, Ankavay M, et al. A genome-wide CRISPR/Cas9 screen identifies a role for Rab5A and early endosomes in hepatitis E virus replication. Proc Natl Acad Sci USA 2023 ; 120 : e2307423120. [Google Scholar]
© 2026 médecine/sciences – Inserm
 Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l’utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l’utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Liste des figures
|
Figure 1 Voies d’entrée des particules eHEV et nHEV dans les hépatocytes. Les particules eHEV et nHEV pénètrent dans les hépatocytes par des voies différentes. La phosphatidylsérine (PS) présente dans l’enveloppe virale de eHEV interagit avec le récepteur TIM1 pour permettre l’entrée du virus. Pour nHEV, différentes protéines de l’hôte, notamment les protéoglycanes à héparane sulfate (HSPG), le récepteur de l’asialoglycoprotéine (ASGPR), et les intégrines, peuvent faciliter son entrée. De plus, le récepteur du facteur de croissance épidermique (EGFR) et la tyrosine kinase YES1 jouent également un rôle dans les étapes d’entrée des deux types de particules. Fu et al. ont montré que eHEV transite probablement par les endosomes précoces (Rab5+) et tardifs (Rab7+) pour atteindre les lysosomes (Lamp1+), tandis que nHEV transite préférentiellement par les endosomes de recyclage (Rab11+) et les endosomes tardifs avant d’atteindre les lysosomes. Dans ce dernier compartiment, la membrane virale de eHEV est dégradée par des lipases et la capside par les cathepsines (Pac-mans verts), puis l’ARN viral est libéré dans le cytoplasme, ce qui permet la transcription et la traduction du génome viral. Figure réalisée avec BioRender. |
| Dans le texte | |
|
Figure 2 Technique d’imagerie RNAscope pour l’ARN de HEV. Le génome du virus de l’hépatite E (HEV) est constitué d’un ARN de 7,2 kilobases (kb) organisé en trois cadres ouverts de lecture (ORF) codant trois protéines virales. ORF1 code une polyprotéine impliquée dans la réplication virale, ORF2 code la protéine de capside, et ORF3 code une petite phosphoprotéine impliquée dans la sécrétion des particules virales. Les protéines ORF2 et ORF3 sont produites à partir d’un ARN sous-génomique de 2,2 kb. RNC : région non codante. Une représentation schématique de la technique RNAscope est présentée. Cette technique d’hybridation in situ « multiplexée » permet la détection spécifique de l’ARN par hybridation de paires de sondes dites « ZZ » sur des séquences cibles adjacentes. Fu et al. ont utilisé deux sondes différentes : une sonde ciblant une région conservée de ORF1, permettant de détecter spécifiquement l’ARN génomique de HEV (flèche en trait pointillé rouge bordeaux ), et une sonde ciblant une région chevauchante de ORF2 et ORF3, permettant de détecter à la fois l’ARN génomique et l’ARN sous-génomique de HEV (flèche en trait pointillé bleu). La combinaison de ces deux sondes permet de distinguer les génomes complets de HEV, encapsidés dans les particules virales, des génomes défectueux, tronqués ou sous-génomiques. Cette distinction est importante pour étudier le processus d’entrée du virus dans la cellule hôte. Après hybridation des paires ZZ, un système d’amplification du signal est assuré par la liaison séquentielle de molécules pré-amplificatrices et amplificatrices, suivie de l’hybridation de multiples sondes fluorescentes, produisant un signal ponctué, spécifique et sensible, correspondant à des molécules individuelles d’ARN. Figure réalisée avec BioRender. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.