")
")
| Issue |
Med Sci (Paris)
Volume 42, Number 5, Mai 2026
|
|
|---|---|---|
| Page(s) | 424 - 426 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/2026073 | |
| Published online | 29 mai 2026 | |
ARHGEF18, un facteur d’échange nucléotidique mécano-sensible contrôlant les jonctions serrées de l’endothélium vasculaire
ARHGEF18 is a flow-responsive nucleotide exchange factor controlling endothelial tight junctions and vascular leakage
Nantes Université, CNRS, Inserm, Institut du thorax, Nantes, France
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
La mécano-transduction des cellules de l’endothélium vasculaire désigne le processus physiologique par lequel ces cellules transforment un stimulus mécanique lié au flux sanguin en un signal intracellulaire de nature biochimique. Dans les vaisseaux sanguins, les cellules endothéliales sont en effet constamment soumises à des forces mécaniques produites par l’écoulement du sang, appelées contraintes de cisaillement. La réponse adaptative de ces cellules aux contraintes de cisaillement contribue à l’homéostasie vasculaire [1]. Une réponse défectueuse contribue en revanche à l’apparition de maladies vasculaires comme l’hypertension artérielle, l’athérosclérose, ou l’anévrisme artériel intracrânien [2, 3].
Les protéines et complexes moléculaires mécano-sensibles présents à la surface des cellules endothéliales jouent un rôle essentiel dans ce processus de transduction, participant ainsi à l’alignement des cellules dans le sens du flux, à la production de monoxyde d’azote, à la libération de chimiokines, et à la réorganisation des jonctions intercellulaires [3]. La réorganisation du cytosquelette d’actine impliquant les petites protéines G de la famille Rho [4] est un phénomène clé dans cette réponse. Le flux sanguin induit le recrutement des protéines Rho à la membrane plasmique et leur activation par les facteurs d’échange nucléotidique des protéines Rho (Rho guanine nucleotide exchange factors, RhoGEF), favorisant leur transition d’un état inactif lié au GDP (guanosine diphosphate) à un état actif lié au GTP (guanosine triphosphate) [5]. Alors que les protéines Rho sont ubiquitaires, l’expression des RhoGEF peut différer entre les organes ou les tissus, et leur activation peut dépendre d’un stimulus ou d’une temporalité spécifique [6], ce qui leur permet de contrôler finement les effets des protéines Rho sur les fonctions cellulaires.
ARHGEF18, un facteur d’échange nucléotidique mécano-sensible
Afin d’identifier les RhoGEF dont la synthèse par les cellules endothéliales est modulée par les contraintes de cisaillement, nous avons réalisé une analyse comparative du transcriptome de ces cellules après les avoir exposées à différents degrés de contrainte : contrainte nulle (0 dyn/cm2), faible (3,6 dyn/cm2, condition pathologique), moyenne (16 dyn/cm2, condition physiologique), ou forte (36 dyn/cm2, condition pathologique). Parmi les transcrits codants des RhoGEF, celui codant la protéine ARHGEF18 (Rho/Rac guanine nucleotide exchange factor 18) était présent en quantité inversement corrélée au degré de la contrainte de cisaillement appliquée, ce qu’une analyse quantitative de ce transcrit par RT-qPCR a confirmé [7]. L’expression d’une Rho-GEF n’étant pas nécessairement corrélée à son degré d’activité [8], nous avons évalué l’activité d’échange nucléotidique de ARHGEF18 sur les protéines RhoA et Rac1 sous ces différentes contraintes de cisaillement. Nous avons ainsi montré que cette activité était restreinte à RhoA, et qu’elle était particulièrement forte lorsque les cellules étaient exposées à des contraintes physiologiques, alors qu’elle diminuait pour les contraintes pathologiques [7].
ARHGEF18 interagit avec les protéines des jonctions intercellulaires serrées
ARHGEF18 est impliquée dans la formation des jonctions serrées entre les cellules épithéliales [9]. Nous avons donc recherché une interaction entre la protéine ARHGEF18 endogène et les protéines des jonctions intercellulaires de l’endothélium. In vitro, nous avons montré, par co-immunoprécipitation, l’interaction de ARHGEF18 avec deux protéines des jonctions serrées, ZO-1 (zonula occludens-1) et la claudine 5, mais pas avec la VE-cadhérine, principal composant des jonctions d’adhérence dans les cellules endothéliales. Par immunofluorescence, nous avons également montré que ARHGEF18 est colocalisée avec ZO-1 [7].
ARHGEF18 contribue à l’adhérence et à la migration des cellules endothéliales in vitro
Pour évaluer le rôle de ARHGEF18 dans les processus cellulaires dépendant de RhoA et de l’organisation du cytosquelette d’actine, nous avons « éteint » l’expression de ARHGEF18 par une stratégie ciblant l’ARN codant pour cette protéine (en utilisant la technique des petits ARN interférents ou celle des petits ARN en épingle à cheveux). Des mesures d’impédance en temps réel ont montré que ARHGEF18 participe à l’adhérence et à l’étalement des cellules endothéliales sur une matrice extracellulaire in vitro. Dans ce système expérimental, l’effet délétère d’une perte de ARHGEF18 implique une diminution des points focaux d’adhérence au support. Enfin, nous avons montré que la perte de ARHGEF18 réduit la migration des cellules endothéliales dans un test de cicatrisation in vitro [7].
L’activité d’échange nucléotidique de ARHGEF18 est impliquée dans la réponse des cellules endothéliales à un flux physiologique
Nous avons ensuite cherché à savoir dans quelle mesure ARHGEF18 contribue à la réponse des cellules endothéliales au flux sanguin. Nous avons montré que la déplétion de ARHGEF18 conduit à un défaut d’élongation et d’alignement de ces cellules dans le sens du flux liquidien in vitro, ainsi qu’à une réduction de la quantité de ZO-1 aux jonctions serrées et de l’actine corticale. À cette déstabilisation des jonctions serrées et de l’actine corticale était associée l’apparition de brèches entre les cellules. Sachant que la mobilité cellulaire participe à l’intégrité de la barrière endothéliale [10] et que les cellules endothéliales rendues déficientes en ARHGEF18 sont moins mobiles que les cellules témoins dans le test de cicatrisation, nous avons également testé la mobilité des cellules privées de ARHGEF18 en présence des contraintes de cisaillement. Nous avons alors constaté, par microscopie en temps réel, que ces cellules se déplaçaient moins vite et moins collectivement que les cellules témoins, ce qui provoquait des tensions intercellulaires conduisant à la rupture de la barrière endothéliale. Étonnamment, cet effet sur la barrière endothéliale n’a pas été constaté en condition « statique », c’est-à-dire en l’absence de flux [7].
L’étape suivante était de déterminer si l’activité d’échange nucléotidique (activité GEF) de ARHGEF18 est nécessaire à la réponse des cellules endothéliales aux contraintes de cisaillement. En utilisant un mutant de ARHGEF18 dépourvu d’activité GEF, nous avons montré que cette perte d’activité empêche l’interaction de ARHGEF18 avec RhoA dans les cellules endothéliales, et qu’elle suffit à provoquer un défaut d’alignement de ces cellules en réponse au flux ainsi qu’un défaut de la localisation de ZO-1 aux jonctions intercellulaires. De plus, l’extinction de l’expression de ARHGEF18 ou l’expression de son mutant dépourvu d’activité GEF conduit à une perte de la phosphorylation de la protéine p38, qui intervient dans des voies de signalisation impliquées dans la mécano-sensibilité et le remodelage du cytosquelette d’actine [11]. L’activité de ARHGEF18 dans ces cellules semble relayée par p38, puisque l’inhibition pharmacologique de cette protéine reproduit les effets de la perte de ARHGEF18 : défaut d’alignement des cellules, délocalisation de ZO-1, et désorganisation des adhérences focales [7]. Pour être active, ARHGEF18 doit être phosphorylée sur le résidu tyrosine 606 de son domaine d’homologie Dbl (DH, ou domaine RhoGEF) qui induit le déplacement du GDP [9]. Dans les cellules endothéliales soumises à des contraintes de cisaillement physiologiques, nous avons montré l’implication de la protéine YES, une kinase de la famille Src, dans ce processus d’activation : son inhibition pharmacologique ou son extinction par la technique d’interférence ARN entraîne une diminution de la phosphorylation de ARHGEF18 sur le résidu tyrosine 606 [7].
ARHGEF18 intervient dans l’angiogenèse et le contrôle de la barrière endothéliale chez la souris
Nous avons produit des souris dépourvues de ARHGEF18 spécifiquement dans les cellules endothéliales. Chez ces souris, six jours après la naissance, le réseau vasculaire de la rétine présentait une expansion radiale et une densité vasculaire réduites. La localisation de ZO-1 aux jonctions intercellulaires de l’endothélium était perturbée dans les artères de la rétine ainsi que dans l’aorte, où les cellules endothéliales apparaissaient également moins bien orientées dans le sens du flux sanguin. Dans la rétine et le cerveau, nous avons constaté, par immunofluorescence, une extravasion d’érythrocytes traduisant une perméabilité accrue de l’endothélium. Cette perméabilité anormale ne concernait pas tout l’organisme puisque le poumon et le foie, par exemple, en étaient préservés. En revanche, les artères rénales et mésentériques étaient affectées elles aussi. Ces constats permettent de conclure à l’implication de ARHGEF18 dans l’angiogenèse physiologique et le contrôle de la barrière endothéliale au cours du développement [7].
Perspectives
En résumé, ARHGEF18 est un RhoGEF mécano-sensible des cellules endothéliales, dont l’activation est dépendante du flux sanguin auquel elles sont soumises. ARHGEF18 participe à la réponse de ces cellules au flux sanguin en favorisant la stabilité des jonctions intercellulaires serrées, l’alignement des cellules endothéliales dans le sens du flux, et la formation du réseau cortical d’actine (Figure 1). L’activation de ARHGEF18 nécessite sa phosphorylation par la kinase YES, et ses effets cellulaires impliquent la voie de signalisation de p38. Enfin, nous avons montré que ARHGEF18 participe au renforcement de la barrière endothéliale in vivo chez la souris. Il conviendra de s’intéresser au rôle de ARHGEF18 dans des contextes pathologiques tels que l’athérosclérose ou l’anévrisme intracrânien, situations dans lesquelles une perméabilité endothéliale accrue favorise le recrutement de cellules de l’inflammation et le remodelage de la paroi des artères.
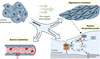 |
Figure 1 Rôle de ARHGEF18 dans le contrôle des jonctions serrées de l’endothélium. A18 : ARHGEF18 ; CLD5 : claudine 5 ; ZO1 : zonula occludens 1. |
Liens d’intérêt
Les auteurs déclarent ne pas avoir de lien d’intérêt.
Références
- Wang C, Baker BM, Chen CS, et al. Endothelial cell sensing of flow direction. Arterioscler Thromb Vasc Biol 2013 ; 33 : 2130-6. [Google Scholar]
- Chalouhi N, Hoh BL, Hasan D. Review of cerebral aneurysm formation, growth, and rupture. Stroke 2013 ; 44 : 3613-22. [Google Scholar]
- Pan S. Molecular mechanisms responsible for the atheroprotective effects of laminar shear stress. Antioxid Redox Signal 2009 ; 11 : 1669-82. [Google Scholar]
- Tzima E. Role of small GTPases in endothelial cytoskeletal dynamics and the shear stress response. Circ Res 2006 ; 98 : 176-85. [Google Scholar]
- Etienne-Manneville S, Hall A. Rho GTPases in cell biology. Nature 2002 ; 420 : 629-35. [Google Scholar]
- Cook DR, Rossman KL, Der CJ. Rho guanine nucleotide exchange factors: regulators of Rho GTPase activity in development and disease. Oncogene 2014 ; 33 : 4021-35. [Google Scholar]
- Batta SPR, Rio M, Lebot C, et al. ARHGEF18 is a flow-responsive exchange factor controlling endothelial tight junctions and vascular leakage. Cell Reports 2025 ; 44 : 115288. [Google Scholar]
- Cario-Toumaniantz C, Ferland-McCollough D, Chadeuf G, et al. RhoA guanine exchange factor expression profile in arteries: evidence for a Rho kinase-dependent negative feedback in angiotensin II-dependent hypertension. Am J Physiol Cell Physiol 2012 ; 302 : C1394-404. [Google Scholar]
- Terry SJ, Zihni C, Elbediwy A, et al. Spatially restricted activation of RhoA signalling at epithelial junctions by p114RhoGEF drives junction formation and morphogenesis. Nat Cell Biol 2011 ; 13 : 159-66. [Google Scholar]
- Szymborska A, Gerhardt H. Hold me, but not too tight: Endothelial cell-cell junctions in angiogenesis. Cold Spring Harb Perspect Biol 2018 ; 10 : a029223. [Google Scholar]
- Vion AC, Perovic T, Petit C, et al. Endothelial cell orientation and polarity are controlled by shear stress and VEGF through distinct signaling pathways. Front Physiol 2020 ; 11 : 623769. [Google Scholar]
© 2026 médecine/sciences – Inserm
 Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l’utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l’utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Liste des figures
|
Figure 1 Rôle de ARHGEF18 dans le contrôle des jonctions serrées de l’endothélium. A18 : ARHGEF18 ; CLD5 : claudine 5 ; ZO1 : zonula occludens 1. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.