")
")
| Issue |
Med Sci (Paris)
Volume 42, Number 5, Mai 2026
|
|
|---|---|---|
| Page(s) | 459 - 466 | |
| Section | M/S Revues | |
| DOI | https://doi.org/10.1051/medsci/2026081 | |
| Published online | 29 mai 2026 | |
Thyroïde et équilibre
Un lien surprenant entre hormones et dysfonctionnements vestibulaires
Thyroid and balance: a surprising link between hormones and vestibular dysfunction
Aix Marseille université-CNRS UMR 7077, Centre de recherche en psychologie et neurosciences (CRPN), Marseille, France
**
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Résumé
Des données récentes montrent que la thyroxine peut accélérer la restauration de l’équilibre après une vestibulopathie unilatérale aiguë. Nous proposons ici une synthèse des interactions entre l’axe thyroïdien et le système vestibulaire, depuis le développement de l’oreille interne jusqu’aux mécanismes cellulaires impliqués dans la compensation. Les effets multiples des hormones thyroïdiennes en font une cible thérapeutique potentiellement prometteuse, bien que les études dans ce domaine soient encore limitées. En approfondissant cette thématique, nous pourrions découvrir de nouvelles perspectives susceptibles de mieux éclairer la compréhension et le traitement des dysfonctionnements vestibulaires. Ce travail ouvre ainsi la voie à des recherches futures qui pourraient non seulement enrichir notre approche clinique, mais aussi améliorer la qualité de vie des patients souffrant de troubles de l’équilibre.
Abstract
Recent data show that thyroxine can accelerate the restoration of balance after acute unilateral vestibular disorder. Here, we present a synthesis of the interactions between the thyroid axis and the vestibular system, from the development of the inner ear to the cellular mechanisms involved in compensation. The diverse effects of thyroid hormones make them a promising therapeutic target, although studies in this area remain limited. By exploring this topic further, we may discover new perspectives for understanding and treating vestibular dysfunctions. This work thus paves the way for future research that could not only improve our clinical approach but also improve outcomes for patients suffering from balance disorders.
Co-premier auteur
© 2026 médecine/sciences – Inserm
 Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l’utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l’utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Avant d’aborder les liens entre désordres vestibulaires1 et dysfonctionnements thyroïdiens, il convient de rappeler brièvement l’organisation et le fonctionnement du système vestibulaire périphérique [1, 2]. Celui-ci est constitué de trois canaux semi-circulaires et de deux organes otolithiques2, l’utricule et le saccule. Les canaux semi-circulaires, disposés selon trois plans orthogonaux, permettent la détection des rotations de la tête grâce au déplacement de l’endolymphe qu’ils contiennent. Les organes otolithiques assurent la détection des accélérations linéaires et de l’orientation de la tête par rapport à la gravité, avec une contribution dominante de l’utricule dans le plan horizontal et du saccule dans le plan vertical (Figure 1). Les organes otolithiques et les ampoules situés à la base des canaux semi-circulaires reposent sur un épithélium sensoriel spécialisé au sein duquel des cellules ciliées mécanosensibles transforment les déformations mécaniques liées aux mouvements de la tête en signaux neuronaux relayés vers le système nerveux central. Toute altération de ces mécanorécepteurs, de leur environnement, des voies de transmission associées ou des structures centrales (noyaux vestibulaires situés dans le tronc cérébral) peut conduire à une dysfonction vestibulaire et à l’apparition de troubles de l’équilibre.
Sur le plan clinique, ces atteintes se traduisent par un syndrome vestibulaire (Figure 2), associant vertiges, instabilité posturale, troubles de la marche, une perte de fixation du regard pendant un déplacement, des mouvements involontaires et rythmiques des yeux (nystagmus) et fréquemment des nausées. Bien que les mécanismes physiopathologiques exacts des maladies vestibulaires restent encore débattus, le vertige positionnel paroxystique bénin3 est généralement associé à un dysfonctionnement des organes otolithiques, consécutif au détachement d’otoconies, également appelés otolithes, qui migrent dans les canaux semi-circulaires, où ils provoquent une stimulation vestibulaire inappropriée lors des changements de position de la tête [3]. La maladie de Ménière4 est traditionnellement reliée à une altération de l’homéostasie de l’endolymphe [4], pouvant entraîner dans certains cas une distension des compartiments endolymphatiques (hydrops endolymphatique), tandis que la vestibulopathie unilatérale aiguë5 est le plus souvent attribuée à une atteinte du nerf vestibulaire ou du labyrinthe [5]. Au-delà des manifestations cliniques, plusieurs travaux récents suggèrent que le fonctionnement du système vestibulaire pourrait être modulé par des facteurs endocriniens, et notamment par les hormones thyroïdiennes, ouvrant ainsi de nouvelles perspectives pour comprendre la physiopathologie du système vestibulaire.
 |
Figure 2 Syndrome vestibulaire et signes pathologiques. Suite à une perte vestibulaire unilatérale, nous observons une quadruple symptomatologie composée de signes oculomoteurs, perceptivo-cognitifs, végétatifs et posturolocomoteurs. Ce syndrome résulte d’un déséquilibre électrophysiologique entre les noyaux vestibulaires droit et gauche, consécutif à la perte d’afférences vestibulaires unilatérales. |
Des désordres vestibulaires peuvent-ils être causés par un dérèglement thyroïdien ? Cette question suscite aujourd’hui un intérêt croissant à la suite de nombreuses données liant l’hypothyroïdie, l’autoimmunité et les troubles de l’équilibre.
Une vaste étude de cohorte coréenne a montré que les antécédents d’hypothyroïdie, d’hyperthyroïdie ou de thyroïdite augmentent le risque de développer une maladie de Ménière, en particulier chez les femmes [6]. De même, Lin et al. ont rapporté que les patients hypothyroïdiens présentent un risque significativement accru de développer une maladie de Ménière par rapport aux sujets euthyroïdiens6 [7]. Ce résultat a également été observé dans une cohorte de patients syriens [8]. Récemment, une méta-analyse d’observations cliniques a indiqué que l’hypothyroïdie aggrave l’instabilité posturale et les troubles de la marche dans les vertiges d’origine centrale ou périphérique [9]. Bien que l’efficacité d’un traitement substitutif à la thyroxine sur les symptômes vertigineux reste controversée [7], certaines études suggèrent une amélioration partielle des troubles de l’équilibre [10,11].
Dans le vertige positionnel paroxystique bénin, le lien avec l’hypothyroïdie reste débattu. Si certaines études ne rapportent aucune association [12–14], d’autres rapportent un risque accru de récidive du vertige positionnel paroxystique bénin chez les patients hypothyroïdiens sous traitement hormonal [15].
D’autres études se sont intéressées plus spécifiquement aux liens entre les vertiges et les hypothyroïdies auto-immunes, telles que la thyroïdite d’Hashimoto7 (pour revue : [16,17]). En effet, les maladies inflammatoires, telles que la maladie d’Hashimoto (inflammation chronique auto-immune de la thyroïde provoquant une hypothyroïdie), pourraient provoquer une réaction immunitaire croisée contre les cellules de l’oreille interne et altérer les fonctions cochléaires et vestibulaires [18]. Fattori et al. ont montré que la prévalence des auto-anticorps anti-thyroïdiens était significativement plus élevée chez les patients atteints de la maladie de Ménière (38 %) que chez les témoins sains (7 %) [19]. Ces données ont été confirmées par d’autres équipes et suggèrent que les patients atteints de la maladie de Ménière ou de vertige positionnel paroxystique bénin sont de potentiels candidats pour le développement d’une thyroïdite de Hashimoto et inversement [20–22].
Le lien étroit entre dysfonctionnement thyroïdienne et troubles vestibulaires pourrait s’expliquer par le rôle essentiel des hormones thyroïdiennes dans le développement de l’oreille interne, où elles régulent la maturation des cellules sensorielles et l’organisation des structures vestibulaires. Chez l’adulte, l’expression des récepteurs aux hormones thyroïdiennes dans les noyaux vestibulaires centraux suggère qu’elles pourraient moduler l’activité des circuits vestibulaires en conditions physiologiques. Une perturbation de cette signalisation endocrine pourrait ainsi contribuer à l’apparition de troubles vestibulaires.
Les hormones thyroïdiennes dans le développement de l’oreille interne
Au-delà du rôle des hormones thyroïdiennes dans le développement cérébral, ces hormones participent également au développement de l’oreille interne. L’hypothyroïdie congénitale entraîne un retard dans le développement des systèmes auditif et vestibulaire [23]. Si de nombreuses études se sont intéressées aux altérations auditives liées à ce trouble [24–27], les conséquences sur les fonctions vestibulaires demeurent beaucoup moins explorées [28–30]. Chez des rats nouveau-nés atteints d’hypothyroïdie congénitale, Meza et ses collaborateurs ont démontré l’absence de nystagmus8 post-rotatoire après stimulation vestibulaire, confirmant leurs travaux antérieurs montrant l’absence de réponse des canaux semi-circulaires chez ces mêmes animaux [29,31]. Sur le plan cellulaire, l’hypothyroïdie congénitale induit un retard de différenciation des cellules ciliées vestibulaires, ainsi qu’un défaut de formation des synapses en calice et de myélinisation des fibres vestibulaires primaires [28]. Les hormones thyroïdiennes jouent donc un rôle essentiel dans le développement de l’appareil vestibulaire périphérique. Plus récemment, il a été montré que leurs récepteurs ainsi que l’enzyme responsable de la conversion de la thyroxine (T4) en sa forme active, la triiodothyronine (T3), sont également présents dans les noyaux vestibulaires de rats adultes [32]. Le rôle de ces récepteurs au sein des structures vestibulaires centrales demeurait toutefois inconnu. C’est dans ce contexte que notre équipe de recherche a entrepris d’en étudier les fonctions, en particulier leurs implications potentielles dans les processus de compensation vestibulaire consécutifs à une lésion périphérique du système vestibulaire.
Démonstration de l’effet bénéfique d’un traitement aigu à la thyroxine sur la compensation vestibulaire
Depuis plusieurs années, notre équipe explore des approches pharmacologiques destinées à réduire le syndrome vestibulaire et à favoriser la restauration fonctionnelle dans des modèles animaux de pathologies de l’oreille interne. Chez le têtard, il a été montré que l’ablation chirurgicale de l’hypophyse, qui contrôle l’activité de la thyroïde9, retarde la compensation vestibulaire après labyrinthectomie10 unilatérale, alors que des injections de thyroxine restaurent la cinétique de compensation chez ces animaux dépourvus d’hypophyse [33]. De même, une perfusion sous-cutanée continue de TRH (thyrotropin-releasing hormone, l’hormone hypothalamique responsable de la libération de T4 et T3) pendant 14 jours accélère la compensation vestibulaire chez des singes adultes soumis à une labyrinthectomie unilatérale [34].
Dans notre étude, nous avons réalisé une section unilatérale du nerf vestibulocochléaire chez des rats adultes afin d’induire un syndrome vestibulaire. Pendant les trois premiers jours suivant la chirurgie, un groupe a reçu des injections intrapéritonéales d’hormone T4 (10 µg/kg), l’autre groupe une solution saline [32]. Comme décrit précédemment, cette dose induit un état transitoire d’hyperthyroïdie, qui revient à la normale en une semaine [35]. Nous avons montré que ce traitement réduit d’environ 30 % les déficits vestibulaires dès le premier jour. Il accélère également la récupération des fonctions posturales et locomotrices altérées par la lésion. Par exemple, la surface de support d’appui au sol, un indicateur fiable de la stabilité posturale, retrouvait sa valeur normale dès le deuxième jour post-lésion chez les rats traités, contre quatorze jours chez les animaux non traités. Une synthèse des résultats principaux de cette étude est présentée dans la Figure 3. Une section unilatérale du nerf vestibulocochléaire reproduit une déafférentation11 soudaine, complète et irréversible des capteurs sensoriels vestibulaires. Cette intervention chirurgicale imite la neurotomie12 pratiquée chez les patients atteints de maladie de Ménière réfractaires aux traitements médicamenteux et permet d’étudier les conséquences cliniques de la perte totale unilatérale d’une entrée vestibulaire. D’autres modèles, tels que la lésion excitotoxique13, permettent d’étudier des atteintes vestibulaires moins sévères et réversibles [36]. C’est le cas de l’administration transtympanique d’acide kaïnique14, un agoniste des récepteurs du glutamate, qui provoque une excitotoxicité et une déafférentation transitoire des cellules ciliées, accompagnée de déficits posturolocomoteurs et vestibulo-oculaires plus légers que ceux observés dans le modèle de section du nerf vestibulaire. Nous avons utilisé ce modèle chez le rat adulte dans l’objectif d’évaluer l’effet anti-vertigineux des hormones thyroïdiennes T4 et T3, ainsi que du métabolite bioactif de l’hormone thyroïdienne, l’acide triiodothyroacétique (TRIAC). Dans ce modèle moins sévère, aucun effet bénéfique des hormones thyroïdiennes n’a été observé [37].
 |
Figure 3 Amélioration des fonctions vestibulaires par un traitement hormonal (T4). Chez les rats ayant reçu de la T4 (10 µg/kg pendant 3 jours), les troubles vestibulaires sont significativement atténués dès le premier jour post-lésion (réduction d’environ 30 % du score des déficits). La stabilité posturale, évaluée par la surface de support au sol (trait jaune), est restaurée dès le deuxième jour, tandis que l’analyse de la locomotion par suivi vidéo révèle une récupération précoce des déplacements chez les animaux traités. Ces effets comportementaux sont associés à une augmentation du nombre de neurones positifs à la cytochrome oxydase dans le noyau vestibulaire latéral du côté lésé, suggérant une implication du métabolisme énergétique neuronal par le traitement à l’hormone T4. |
La différence d’efficacité de l’hormone T4 entre les deux modèles interroge les mécanismes et les cibles d’action des hormones thyroïdiennes : pourquoi l’hormone T4 améliore-t-elle les déficits dans le modèle de section du nerf définitif, mais pas dans celui de lésion excitotoxique réversible ? La glie pourrait expliquer cette différence de réponse à la T4. Lors d’une section du nerf cochléovestibulaire, une forte réaction gliale [une augmentation significative du nombre de cellules GFAP-positives (astrocytes) et IBA1+ (microglies)] se déclenche dans les noyaux vestibulaires désafférentés alors qu’elle est absente dans le modèle excitotoxique [38,39]. Or, les astrocytes expriment l’enzyme D2 (iodothyronine désiodase de type 2), capable de convertir la T4 en T3, forme active des hormones thyroïdiennes qui agit ensuite sur les neurones via ses récepteurs spécifiques (Figure 4). Cette conversion locale, amplifiée par une glie réactive, pourrait renforcer la disponibilité en T3 dans les noyaux vestibulaires et ainsi favoriser les effets des hormones thyroïdiennes sur la compensation vestibulaire. Dans ce contexte, il est tentant de proposer que l’activation gliale, en interaction avec les hormones thyroïdiennes apportées par le traitement, favorise également la plasticité synaptique des réseaux vestibulaire. Une synaptogenèse centrale n’a été décrite, chez le rat adulte, que dans des modèles de section du nerf vestibulaire [40], caractérisés par une déafférentation irréversible et une forte activation gliale. À l’inverse, les modèles de lésions excitotoxiques par l’acide kaïnique, plus modérés et réversibles, ne s’accompagnent pas de synaptogenèse centrale. Cette dissociation entre les deux modèles renforce l’hypothèse selon laquelle l’activation gliale constitue un prérequis à l’engagement de mécanismes de plasticité synaptique centrale. Dans ce cadre, la conversion astrocytaire de T4 en T3 pourrait renforcer localement les processus de plasticité synaptique [40–43], tandis que la microglie activée agirait comme un régulateur clé du remodelage synaptique, facilitant la compensation vestibulaire.
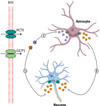 |
Figure 4 Modèle proposé de l’interaction astrocyte-neurone et du transport de la T3 dans le cerveau. Tout d’abord l’hormone T4 (et à moindre mesure l’hormone T3) traverse la barrière hémato-encéphalique (BHE) grâce aux transporteurs OATP1 (organic anion transporting polypeptide 1) et MCT8 (monocarboxylate transporter 8). Trois scénarios sont possibles pour la suite : 1. L’hormone T4 est transportée dans les astrocytes par un transporteur encore inconnu. Dans les astrocytes, l’hormone T4 est convertie en T3 par l’enzyme D2. L’hormone T3 biologiquement active est ensuite transportée hors des astrocytes par un autre transporteur inconnu. 1b. L’hormone T3 depuis la BHE rejoint les neurones par MCT8. 2. L’assimilation neuronale de l’hormone T4 métabolisée en T3 par les astrocytes est facilitée par MCT8 puis l’hormone T3 exerce son action génomique en se liant à son récepteur nucléaire (TR). |
Quand les hormones thyroïdiennes stimulent la plasticité vestibulaire
Si la présence de récepteurs aux hormones thyroïdiennes dans les noyaux vestibulaires suggère une implication potentielle de ces hormones dans le fonctionnement vestibulaire, le rôle physiologique de cette signalisation, en conditions normales, demeure largement méconnu. Dans un contexte pathologique tel que la déafférentation vestibulaire, cette voie pourrait néanmoins être localement activée ou amplifiée, favorisant la restauration de l’équilibre fonctionnel. Néanmoins, le rôle pléiotrope de ces hormones, dont les récepteurs sont exprimés dans divers organes et tissus, suggère que leurs effets pourraient également résulter de mécanismes périphériques ou systémiques. Ainsi, les hormones thyroïdiennes pourraient agir à plusieurs niveaux, modulant non seulement l’excitabilité neuronale et le métabolisme énergétique des neurones vestibulaires, mais également en influençant le tonus musculaire et le métabolisme général, contribuant globalement à la restauration des fonctions vestibulaires.
L’excitabilité neuronale
Après une lésion vestibulaire périphérique, la perte des informations en provenance de l’oreille interne entraîne une perte rapide et importante de l’activité neuronale dans les noyaux vestibulaires ipsilatéraux15 à la lésion. Cette réduction de l’activité perturbe l’équilibre fonctionnel du système vestibulaire, à l’origine des déficits du syndrome vestibulaire. La restauration d’une excitabilité neuronale dans le réseau vestibulaire désafférenté est donc un facteur clé de la compensation vestibulaire [44–47]. Or, plusieurs études suggèrent que les hormones thyroïdiennes modulent l’excitabilité neuronale. Par exemple, des doses élevées des hormones T3 ou T4 réduisent les courants inhibiteurs GABAergiques dans des neurones hippocampiques en culture [48,49], tandis que des traitements par l’hormone T4 in vivo augmentent la fréquence de décharge spontanée dans diverses régions cérébrales, notamment le bulbe olfactif [50] et le gyrus denté16 [51].
Une étude récente a également mis en évidence que l’hormone T3 chez la souris modifie la transmission synaptique excitatrice et inhibitrice [52]. Les mécanismes sous-jacents, combinant potentiellement les effets génomiques et non génomiques des hormones thyroïdiennes, restent toutefois à préciser. En effet, les mécanismes génomiques résultent de l’action des hormones thyroïdiennes avec leurs récepteurs nucléaires, pour moduler l’expression de différents gènes, tandis que les effets non génomiques médiés par des récepteurs membranaires ou cytoplasmiques impliquent des mécanismes rapides indépendants de la modulation de l’expression des gènes. Dans notre modèle, l’effet précoce du traitement sur la compensation vestibulaire suggère une action rapide, probablement impliquant ces deux modes d’action. Si leur action sur l’excitabilité neuronale constitue un premier levier sur la compensation vestibulaire, les hormones thyroïdiennes exercent une influence majeure sur le métabolisme énergétique que nous avons exploré pour éclairer leur impact sur la récupération fonctionnelle après une atteinte vestibulaire.
Le métabolisme énergétique
Les hormones thyroïdiennes occupent une place centrale dans le métabolisme cellulaire. Elles régulent de multiples voies de signalisation impliquées dans le métabolisme des glucides, des lipides et des protéines au sein de nombreux tissus cibles ainsi que la respiration cellulaire mitochondriale [53,54]. Dans nos travaux, nous avons montré qu’un traitement par l’hormone T4 induit une augmentation du nombre de neurones exprimant l’enzyme cytochrome oxydase dans le noyau vestibulaire latéral ipsilatéral à la lésion [32]. L’augmentation de cette enzyme clé du cycle de Krebs, impliquée dans la production d’ATP, suggère que l’hormone T4 stimule la production énergétique neuronale au sein des noyaux vestibulaires. Cet effet favoriserait la restauration de l’excitabilité dans les noyaux vestibulaires lésés, contribuant à la compensation fonctionnelle. Dans cette même logique d’augmentation du métabolisme énergétique neuronal, nous avons observé qu’un traitement par l’hormone T4 prévenait la diminution de l’expression de la protéine KCC2 (K+-Cl- cotransporteur 2), le régulateur majeur de l’homéostasie du chlorure et de l’inhibition neuronale, habituellement observée dans notre modèle de section du nerf vestibulaire. Ce résultat corrobore l’hypothèse de Kaila et al., selon laquelle la diminution de l’expression de KCC2 après une lésion neuronale représenterait une réponse adaptative visant à réduire la dépense énergétique nécessaire au maintien d’un faible gradient intracellulaire de chlorure [55]. L’activation métabolique induite par l’hormone T4 réactiverait donc ce mécanisme compensatoire visant à économiser l’énergie cellulaire non utilisée.
Parallèlement, l’augmentation du métabolisme énergétique induite par les hormones thyroïdiennes s’accompagne d’une augmentation de l’activité locomotrice (distance parcourue) et exploratoire [52], comme nous l’avons également observé dans notre étude [32]. Bien que les effets bénéfiques de la rééducation vestibulaire et de l’exercice physique sur la récupération aient été largement démontrés [56–58], la réduction de l’activité physique est fréquemment observée chez les patients et est associée à une compensation incomplète et à la persistance de symptômes chroniques [59]. Chez des patients opérés d’un schwannome17 vestibulaire, une activité physique régulière avant l’intervention favorise la récupération posturale et locomotrice [60]. Ainsi, en stimulant le métabolisme énergétique et la motricité, les hormones thyroïdiennes pourraient agir comme des catalyseurs de la récupération fonctionnelle après une atteinte vestibulaire.
Le tonus musculaire
Un dernier aspect important à considérer concerne l’action des hormones thyroïdiennes et leur influence sur les muscles squelettiques. Dans la continuité de la section précédente, il est bien établi que les exercices de marche et l’activité physique sont recommandés par les ORL (otorhinolaryngologistes) spécialistes des vertiges. Outre la diminution de l’activité physique souvent observée après une atteinte vestibulaire unilatérale, les premiers jours sont marqués par une nette asymétrie posturale [61]. Dans les modèles animaux de lésion vestibulaire unilatérale, cette asymétrie reflète une hypotonie du côté ipsilatéral du corps, associée à une baisse de l’activité des muscles extenseurs du côté de la lésion et, en miroir, une hypertonie musculaire controlatérale [62,63].
Cette dimension musculaire ne doit pas être négligée, d’autant que les patients souffrant de troubles vestibulaires chroniques présentent fréquemment une diminution de la masse musculaire ou une sarcopénie18 [64]. Par exemple, la sarcopénie (caractérisée par une faible masse musculaire) associée à l’obésité augmentent considérablement le risque de développer un vertige positionnel paroxystique bénin [65]. Le muscle représente donc une cible essentielle pour compenser les déficits posturaux induits par la perte vestibulaire et les hormones thyroïdiennes pourraient y contribuer. Le muscle squelettique est un tissu hautement sensible aux hormones thyroïdiennes [66]. Les hormones thyroïdiennes modulent 1) la fonction contractile en accélérant les cycles contraction-relaxation, 2) le métabolisme grâce à la stimulation de la biogenèse mitochondriale, et 3) la myogenèse et la régénération musculaire [67,68]. Leur présence dans les muscles extra-oculaires et antigravitaires, tels que le soléaire19 [69], suggère également une action possible sur la stabilisation du regard et le maintien du tonus postural.
Bien qu’aucune étude n’ait encore démontré directement une augmentation du tonus musculaire après administration d’hormones thyroïdiennes, il est possible qu’en renforçant la fonction contractile et le métabolisme musculaire, celles-ci contribuent au rééquilibrage du tonus musculaire après déafférentation vestibulaire, soutenant ainsi la restauration de l’équilibre statique et dynamique.
Conclusion
L’ensemble de ces travaux suggère que les hormones thyroïdiennes pourraient jouer un rôle de modulateur essentiel, bien qu’encore insuffisamment exploré, dans les processus de compensation vestibulaire, selon des mécanismes qui restent à préciser. Au-delà de leurs actions périphériques et centrales, les interactions entre le système vestibulaire et l’axe hypothalamo-hypophysaire ouvrent de nouvelles perspectives impliquant d’autres voies hormonales. Le système vestibulaire constitue le principal capteur sensoriel de la gravité, intégrant en permanence les accélérations linéaires et angulaires afin d’assurer l’orientation spatiale, le contrôle postural et la stabilisation du regard. Toute modification du champ gravitaire, comme celles observées en microgravité ou en hypergravité, entraîne une altération profonde de l’activité vestibulaire et des processus d’adaptation centrale qui en découlent. Dans ce contexte, l’existence de voies neuroanatomiques reliant les noyaux vestibulaires à l’axe du stress (axe hypothalamo-hypophysaire) [70, 71], ainsi que les modifications profondes du système thyroïdien observées en microgravité [72–75] ou en hypergravité [76], souligne l’importance du facteur gravitationnel dans la régulation endocrinienne de ces hormones [77]. Ces observations invitent à reconsidérer le rôle du système vestibulaire et son lien avec l’axe thyréotrope, tout en explorant l’impact des hormones thyroïdiennes sur les processus d’adaptation après une lésion. L’interaction entre la gravité, la fonction thyroïdienne et la plasticité vestibulaire ouvre ainsi un champ de recherche particulièrement prometteur, dont les mécanismes restent encore largement à élucider. Cette nouvelle perspective pourrait non seulement enrichir notre compréhension des mécanismes sous-jacents, mais aussi ouvrir la voie à de nouvelles approches thérapeutiques pour le traitement des troubles vestibulaires.
Liens d’intérêt
Les auteurs déclarent ne pas avoir de lien d’intérêt.
Références
- Spratley J, Marques P, Alexandre P. Chapter 2: The physiology of the vestibular system. Disorders of the vestibular system. Switzerland : Springer, 2023 : 13–26. [Google Scholar]
- Monsanto R da C, Pauna HF, Cureoglu S. The anatomy of the vestibular system. Disorders of the vestibular system. Switzerland : Springer, 2023 : 1–12. [Google Scholar]
- Kao WTK, Parnes LS, Chole RA. Otoconia and otolithic membrane fragments within the posterior semicircular canal in BPPV. Laryngoscope 2017 ; 127 : 709–14. [Google Scholar]
- Attyé A, Eliezer M, Medici M, et al. In vivo imaging of saccular hydrops in humans reflects sensorineural hearing loss rather than Meniere’s disease symptoms. Eur Radiol 2018 ; 28 : 2916–22. [Google Scholar]
- Hegemann SCA, Wenzel A. Diagnosis and treatment of vestibular neuritis/neuronitis or peripheral vestibulopathy (PVP)? Open questions and possible answers. Otol Neurotol 2017 ; 38 : 626–31. [Google Scholar]
- Kim SY, Song YS, Wee JH, et al. Association between Ménière’s disease and thyroid diseases: a nested case-control study. Sci Rep 2020 ; 10 : 18224. [Google Scholar]
- Lin WL, Chen CY, Hsu TY, et al. Hypothyroidism is an independent risk factor for Menière’s disease: A population-based cohort study. Medicine (Baltimore) 2019 ; 98 : e15166. [Google Scholar]
- Bakdounes A, Akashe N, Alchallah MO, et al. Prevalence of Ménière’s disease in syrian patients with hypothyroidism: cross-sectional study. Ann Med Surg 2022 ; 81 : 104405. [Google Scholar]
- Bougerolle V, El Khiati R, El Ahmadi A, et al. Statistical associations between vestibular pathologies and hypothyroidism: a retrospective study. J Clin Med 2024 ; 13 : 1099. [Google Scholar]
- Powers WH. Metabolic aspects of Ménière’s disease. Laryngoscope 1978 ; 88 : 122–9. [Google Scholar]
- Santosh UP, Rao MSS. Incidence of hypothyroidism in Meniere’s disease. JCDR 2016 ; 10 : MC01–03. [Google Scholar]
- Sari K, Yildirim T, Borekci H, et al. The relationship between benign paroxysmal positional vertigo and thyroid autoimmunity. Acta Otolaryngol 2015 ; 135 : 754–7. [Google Scholar]
- Miś kiewicz-Orczyk K, Vlaykov A, Lisowska G, et al. Does thyroid hormone metabolism correlate with the objective assessment of the vestibular organ in patients with vertigo? J Clin Med 2022 ; 11 : 6771. [Google Scholar]
- Lima CM, Paiva DFF, Corona AP, et al. Association between benign paroxysmal positional vertigo and thyroid diseases: systematic review and meta-analysis. Int Arch Otorhinolaryngol 2024 ; 28 : e530–6. [Google Scholar]
- Tricarico L, Di Cesare T, Galli J, et al. Benign paroxysmal positional vertigo: is hypothyroidism a risk factor for recurrence? Acta Otorhinolaryngol Ital 2022 ; 42 : 465–70. [Google Scholar]
- Miś kiewicz-Orczyk KA, Lisowska G, Kajdaniuk D, et al. Can Hashimoto’s thyroiditis cause vertigo? Endokrynol Pol 2020 ; 70 : 76–86. [Google Scholar]
- Chen JJ, Hsu CW, Chen TY, et al. Audiovestibular dysfunction in patients with Hashimoto’s disease: a systematic review. Int J Mol Sci 2025 ; 26 : 4703. [Google Scholar]
- Kim SH, Kim JY, Lee HJ, et al. Autoimmunity as a candidate for the etiopathogenesis of Meniere’s disease: detection of autoimmune reactions and diagnostic biomarker candidate. PloS One 2014 ; 9 : e111039. [Google Scholar]
- Fattori B, Nacci A, Dardano A, et al. Possible association between thyroid autoimmunity and Menière’s disease. Clin Exp Immunol 2008 ; 152 : 28–32. [Google Scholar]
- Chiarella G, Tognini S, Nacci A, et al. Vestibular disorders in euthyroid patients with Hashimoto’s thyroiditis: role of thyroid autoimmunity. Clin Endocrinol 2014 ; 81 : 600–65. [Google Scholar]
- Chiarella G, Russo D, Monzani F, et al. HASHIMOTO thyroiditis and vestibular dysfunction. Endocr Pract 2017 ; 23 : 863–8. [Google Scholar]
- Papi G, Guidetti G, Corsello SM, et al. The association between benign paroxysmal positional vertigo and autoimmune chronic thyroiditis is not related to thyroid status. Thyroid 2010 ; 20 : 237–8. [Google Scholar]
- Rybak L. Metabolic disorders of the vestibular system. Otolaryngol Head Neck Surg 1995 ; 112 : 128–32. [Google Scholar]
- Dror AA, Lenz DR, Shivatzki S, et al. Atrophic thyroid follicles and inner ear defects reminiscent of cochlear hypothyroidism in Slc26a4-related deafness. Mamm Genome 2014 ; 25 : 304–16. [Google Scholar]
- Andrade CLO, Alves C de AD, Ramos HE. Congenital hypothyroidism and the deleterious effects on auditory function and language skills: a narrative review. Front Endocrinol 2021 ; 12 : 671784. [Google Scholar]
- Zhang K, Fried J, Nguyen SA, et al. Dysfunction of the auditory system in children with hypothyroidism: a systematic review and meta-analysis. Ear Hear 2022 ; 43 : 23–31. [Google Scholar]
- Tan J, Wang X, Li Y, et al. Impaired tectorial membrane and ribbon synapse maturation in the cochlea of mice with congenital hypothyroidism. Biochem Biophys Res Commun 2023 ; 655 : 68–74. [Google Scholar]
- Dememes D, Dechesne C, Legrand C, et al. Effects of hypothyroidism on postnatal development in the peripheral vestibular system. Dev Brain Res 1986 ; 25 : 147–52. [Google Scholar]
- Meza G, Acuña D, Escobar C. Development of vestibular and auditory function: Effects of hypothyroidism and thyroxine replacement therapy on nystagmus and auditory evoked potentials in the pigmented rat. Int J Dev Neurosci 1996 ; 14 : 515–22. [Google Scholar]
- Meza G, Acuña D, Gutiérrez A, et al. Development of vestibular function: biochemical, morphological and electronystagmographical assessment in the rat. Int J Dev Neurosci 1996 ; 14 : 507–13. [Google Scholar]
- Meza G, Acuña D, Peñaloza Y, et al. Congenital hypothyroidism. Vestibular and auditory damage in the pigmented rat. Ann NY Acad Sci 1991 ; 630 : 274–6. [Google Scholar]
- Rastoldo G, Marouane E, El-Mahmoudi N, et al. L-thyroxine improves vestibular compensation in a rat model of acute peripheral vestibulopathy: cellular and behavioral aspects. Cells 2022 ; 11 : 684. [Google Scholar]
- Horn E, Rayer B. A hormonal component in central vestibular compensation. Z. Naturforsch 1980 ; 35 : 1120–1. [Google Scholar]
- Ishii M, Igarashi M. Effect of thyrotropin-releasing hormone on vestibular compensation in primates. Am J Otolaryngol 1986 ; 7 : 177–80. [Google Scholar]
- Yu D, Zhou H, Yang Y, et al. The bidirectional effects of hypothyroidism and hyperthyroidism on anxiety- and depression-like behaviors in rats. Horm Behav 2015 ; 69 : 106–15. [Google Scholar]
- Tighilet B, Trico J, Xavier F, et al. What predictability for animal models of peripheral vestibular disorders? Biomedicines 2022 ; 10 : 3097. [Google Scholar]
- Bringuier CM, Hatat B, Boularand R, et al. Characterization of thyroid hormones antivertigo effects in a rat model of excitotoxically-induced vestibulopathy. Front Neurol 2022 ; 13 : 877319. [Google Scholar]
- Campos-Torres A, Touret M, Vidal PP, et al. The differential response of astrocytes within the vestibular and cochlear nuclei following unilateral labyrinthectomy or vestibular afferent activity blockade by transtympanic tetrodotoxin injection in the rat. Neuroscience 2005 ; 130 : 853–65. [Google Scholar]
- Dutheil S, Lacour M, Tighilet B. Neurogenic potential of the vestibular nuclei and behavioural recovery time course in the adult cat are governed by the nature of the vestibular damage. PLoS One 2011 ; 6 : e22262. [Google Scholar]
- Li H, Dokas LA, Godfrey DA, et al. Remodeling of synaptic connections in the deafferented vestibular nuclear complex. J Vestib Res Equilib Orientat 2002 ; 12 : 167–83. [Google Scholar]
- Zhang YX, Li LY, Xing Y, et al. Glutamatergic synaptic plasticity in medial vestibular nuclei during vestibular compensation. Neuroscience 2025 ; 576 : 213–22. [Google Scholar]
- Raymond J, Ez-Zaher L, Demêmes D, et al. Quantification of synaptic density changes in the medial vestibular nucleus of the cat following vestibular neurectomy. Restor Neurol Neurosci 1991 ; 3 : 197–203. [Google Scholar]
- Chang MY, Park S, Choi JJ, et al. MicroRNAs 218a-5p, 219a-5p, and 221-3p regulate vestibular compensation. Sci Rep 2017 ; 7 : 8701. [Google Scholar]
- Cameron SA, Dutia MB. Cellular basis of vestibular compensation: changes in intrinsic excitability of MVN neurones. Neuroreport 1997 ; 8 : 2595–9. [Google Scholar]
- Curthoys IS. Vestibular compensation and substitution. Curr Opin Neurol 2000 ; 13 : 27. [Google Scholar]
- Ris L, Capron B, Vibert N, et al. Modification of the pacemaker activity of vestibular neurons in brainstem slices during vestibular compensation in the guinea pig. Eur J Neurosci 2001 ; 13 : 2234–40. [Google Scholar]
- Beraneck M, Idoux E. Reconsidering the role of neuronal intrinsic properties and neuromodulation in vestibular homeostasis. Front Neurol 2012 ; 3 : 25. [Google Scholar]
- Martin JV, Williams DB, Fitzgerald RM, et al. Thyroid hormonal modulation of the binding and activity of the GABAA receptor complex of brain. Neuroscience 1996 ; 73 : 705–13. [Google Scholar]
- Puia G, Losi G. Thyroid hormones modulate GABAA receptor-mediated currents in hippocampal neurons. Neuropharmacology 2011 ; 60 : 1254–61. [Google Scholar]
- Oshima K, Gorbman A. Influence of thyroxine and steroid hormones on spontaneous and evoked unitary activity in the olfactory bulb of goldfish. Gen Comp Endocrinol 1966 ; 7 : 482–91. [Google Scholar]
- Shabani S, Sarkaki A, Ali Mard S, et al. Central and peripheral administrations of levothyroxine improved memory performance and amplified brain electrical activity in the rat model of Alzheimer’s disease. Neuropeptides 2016 ; 59 : 111–6. [Google Scholar]
- Hochbaum DR, Hulshof L, Urke A, et al. Thyroid hormone remodels cortex to coordinate body-wide metabolism and exploration. Cell 2024 ; 187 : 5679–97.e23. [Google Scholar]
- Mullur R, Liu Y-Y, Brent GA. Thyroid hormone regulation of metabolism. Physiol Rev 2014 ; 94 : 355–82. [Google Scholar]
- Cicatiello AG, Di Girolamo D, Dentice M. Metabolic effects of the intracellular regulation of thyroid hormone: old players, new concepts. Front Endocrinol 2018 ; 9 : 474. [Google Scholar]
- Kaila K, Price TJ, Payne JA, et al. Cation-chloride cotransporters in neuronal development, plasticity and disease. Nat Rev Neurosci 2014 ; 15 : 637–54. [Google Scholar]
- McDonnell MN, Hillier SL. Vestibular rehabilitation for unilateral peripheral vestibular dysfunction. Cochrane Database Syst Rev 2015 ; 1 : CD005397. [Google Scholar]
- Kundakci B, Sultana A, Taylor AJ, et al. The effectiveness of exercise-based vestibular rehabilitation in adult patients with chronic dizziness: a systematic review. F1000Res 2018 ; 7 : 276. [Google Scholar]
- Hall CD, Herdman SJ, Whitney SL, et al. Vestibular rehabilitation for peripheral vestibular hypofunction: an updated clinical practice guideline from the academy of neurologic physical therapy of the american physical therapy association. J Neurol Phys Ther 2022 ; 46 : 118–77. [Google Scholar]
- Van Laer L, Herssens N, Hallemans A, et al. The association between physical activity and chronic symptoms after a unilateral vestibular deafferentation: narrative review. B-ENT 2023 ; 19 : 50–8. [Google Scholar]
- Gauchard GC, Parietti-Winkler C, Lion A, et al. Impact of pre-operative regular physical activity on balance control compensation after vestibular schwannoma surgery. Gait Posture 2013 ; 37 : 82–7. [Google Scholar]
- Strupp M, Bisdorff A, Furman J, et al. Acute unilateral vestibulopathy/vestibular neuritis: Diagnostic criteria: Consensus document of the committee for the classification of vestibular disorders of the Bárány Society. J Vestib Res 2022 ; 32 : 389–406. [Google Scholar]
- Kasri M, Picquet F, Falempin M. Effects of unilateral and bilateral labyrinthectomy on rat postural muscle properties: the soleus. Exp Neurol 2004 ; 185 : 143–53. [Google Scholar]
- Rastoldo G, Marouane E, El Mahmoudi N, et al. Quantitative evaluation of a new posturo-locomotor phenotype in a rodent model of acute unilateral vestibulopathy. Front Neurol 2020 ; 11 : 505. [Google Scholar]
- Jeong S-H, Kwon E. Understanding body composition in vestibular disorders: where do we stand? A narrative review. Res Vestib Sci 2025 ; 24 : 89–97. [Google Scholar]
- Demircan SK, Öner F. The association of sarcopenia and body composition with benign positional paroxysmal vertigo in older adults. Laryngoscope 2025 ; 135 : 1486–92. [Google Scholar]
- Machado M, Bachini F, Itaborahy A. Thyroid hormones and skeletal muscle beyond thermogenesis. J Sci Sport Exerc 2024 ; 6 : 315–23. [Google Scholar]
- Salvatore D, Simonides WS, Dentice M, et al. Thyroid hormones and skeletal muscle — new insights and potential implications. Nat Rev Endocrinol 2014 ; 10 : 206–14. [Google Scholar]
- Bloise FF, Cordeiro A, Ortiga-Carvalho TM. Role of thyroid hormone in skeletal muscle physiology. J Endocrinol 2018 ; 236 : R57–68. [Google Scholar]
- Schmidt ED, Schmidt ED, Gaag R van der, et al. Distribution of the nuclear thyroid-hormone receptor in extraocular and skeletal muscles. J Endocrinol 1992 ; 133 : 67–74. [Google Scholar]
- Saman Y, Bamiou DE, Gleeson M, et al. Interactions between Stress and Vestibular Compensation – A review. Front Neurol 2012 ; 3 : 116. [Google Scholar]
- Saman Y, Arshad Q, Dutia M, et al. Stress and the vestibular system. Int Rev Neurobiol 2020 ; 152 : 221–36. [Google Scholar]
- Plakhuta-Plakutina GI, Kabitskiĭ EN, Dmitrieva NP, et al. Studies of the morphology of the thyroid gland and thyroid hormone levels in the blood of rats in experiments on “Kosmos-1667” and “Kosmos-1887”. Kosm Biol Aviakosm Med 1990 ; 24 : 25–7. [Google Scholar]
- Loginov VI. The inhibition of the thyroid and calcitonin-producing functions of the rat thyroid gland in weightlessness. Aviakosmicheskaia Ekol Meditsina Aerosp Environ Med 1999 ; 33 : 12–6. [Google Scholar]
- Strollo F. Hormonal changes in humans during spaceflight. Adv Space Biol Med 1999 ; 7 : 99–129. [Google Scholar]
- Grimm D, Kossmehl P, Shakibaei M, et al. Effects of simulated microgravity on thyroid carcinoma cells. J Gravit Physiol 2002 ; 9 : 253–6. [Google Scholar]
- Krasnov IB, Alekseev EI, Loginov VI. Role of the endocrine glands in divergence of plastic processes and energy metabolism in rats after extended exposure to hypergravity: cytologic investigation. Aviakosmicheskaia Ekol Meditsina Aerosp Environ Med 2006 ; 40 : 29–34. [Google Scholar]
- Albi E, Krüger M, Hemmersbach R, et al. Impact of gravity on thyroid cells. Int J Mol Sci 2017 ; 18 : 972. [Google Scholar]
- Goldberg ME, Walker MF, Hudspeth AJ. Chapter 40: The vestibular system. Principles of neural science. United States : McGraw-Hill Companies, Inc., 2013 : 917–34. [Google Scholar]
Le système vestibulaire est un organe sensoriel situé dans l’oreille interne qui contribue à la sensation de mouvement et à l’équilibre (ndlr).
L’otolithe (othos : oreille ; lithos : pierre) est une petite structure de carbonate de calcium qui se trouve dans le système vestibulaire (saccule ou utricule) de l’oreille interne des vertébrés. Chez l’être humain, il mesure de 1 à 30 µm en longueur (ndlr).
Le vertige positionnel paroxystique bénin est un trouble fréquent provoquant de courts épisodes de vertige (fausse sensation de mouvements ou de tournis) en réponse à des changements de position de la tête, sans perte auditive (ndlr).
La maladie de Ménière se manifeste par des crises de vertiges, des bourdonnements d’oreille et une perte d’audition (surtout des sons graves). Elle a été ainsi nommée d’après le médecin français Prosper Ménière (1799-1862) qui fut le premier à attribuer à l’oreille interne la maladie associant des vertiges paroxystiques et une surdité (ndlr).
Ce syndrome se caractérise par des symptômes intenses comme un vertige rotatoire, des nausées, des vomissements et une instabilité de l’équilibre (ndlr).
C’est à dire présentant un taux normal d’hormones thyroïdiennes (ndlr).
La thyroïdite de Hashimoto est une maladie auto-immune dont les signes sont ceux d’une hypothyroïdie. Elle est caractérisée notamment par la présence d’anticorps anti-thyroperoxydase et par une infiltration lymphoïde de la glande thyroïde (ndlr).
En médecine, le nystagmus désigne un mouvement oscillatoire involontaire des yeux, généralement lié à une perturbation des circuits nerveux qui contrôlent les mouvements oculaires, notamment les connexions entre l’oreille interne (système vestibulaire) et les muscles qui déplacent les yeux (ndlr).
La thyréostimuline (TSH) est une hormone libérée par l’hypophyse qui régule l’activité de la glande thyroïde. Elle stimule la production des hormones thyroïdiennes, dont les taux dans le sang participent en retour au contrôle de sa sécrétion (ndlr).
Une labyrinthectomie est une intervention chirurgicale qui implique l’ablation ou la destruction du labyrinthe, qui est la structure de l’oreille interne responsable de l’équilibre et de l’audition (ndlr).
La désafférentation désigne la perte des informations sensorielles normalement transmises au cerveau par les voies nerveuses afférentes (ndlr).
Intervention chirurgicale qui consiste à sectionner le nerf de l’équilibre en conservant intact le nerf de l’audition (ndlr).
L’excitotoxicité désigne la destruction ou l’altération des neurones provoquée par une stimulation excessive des récepteurs du glutamate, un neurotransmetteur excitateur (ndlr).
L’acide kaïnique est un composé naturel présent dans l’algue Digenea simplex ou l’amanite tuemouches. C’est un analogue du glutamate et peut agir comme un agoniste spécifique des récepteurs kaïnates, un sous-type de la famille des récepteurs du glutamate (ndlr).
Ipsilatéral, en anatomie, signifie qui se situe ou se produit du même côté du corps (ndlr).
La surface du cerveau, appelée cortex cérébral, est très irrégulière, caractérisée par un schéma distinctif de plis ou de bosses, appelés gyri (singulier : gyrus), et de sillons, appelés sulci (singulier : sulcus). Ces gyri et sulci constituent des repères importants pour séparer le cerveau en centres fonctionnels. Le gyrus denté ou gyrus dentelé est un gyrus du lobe limbique du cortex cérébral. Il est situé au-dessus du sillon hippocampique, le long de l’hippocampe. Il est responsable de la formation de la mémoire et de l’apprentissage (ndlr).
Les schwannomes vestibulaires ou appelés également neurinome de l’acoustique sont des tumeurs bénignes de la gaine du nerf cochléo-vestibulaire, responsable de l’audition et l’équilibre.
La sarcopénie est la perte de masse et de force musculaire associée au vieillissement (ndlr).
Le muscle soléaire est un faisceau du muscle triceps sural, muscle appartenant au mollet. Il permet l’extension du pied. Le muscle soléaire maintient l’équilibre en position debout en agissant comme un stabilisateur pour la cheville
Liste des figures
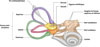 |
Figure 1 Organisation de l’oreille interne. Adapté de [78]. |
| Dans le texte | |
|
Figure 2 Syndrome vestibulaire et signes pathologiques. Suite à une perte vestibulaire unilatérale, nous observons une quadruple symptomatologie composée de signes oculomoteurs, perceptivo-cognitifs, végétatifs et posturolocomoteurs. Ce syndrome résulte d’un déséquilibre électrophysiologique entre les noyaux vestibulaires droit et gauche, consécutif à la perte d’afférences vestibulaires unilatérales. |
| Dans le texte | |
|
Figure 3 Amélioration des fonctions vestibulaires par un traitement hormonal (T4). Chez les rats ayant reçu de la T4 (10 µg/kg pendant 3 jours), les troubles vestibulaires sont significativement atténués dès le premier jour post-lésion (réduction d’environ 30 % du score des déficits). La stabilité posturale, évaluée par la surface de support au sol (trait jaune), est restaurée dès le deuxième jour, tandis que l’analyse de la locomotion par suivi vidéo révèle une récupération précoce des déplacements chez les animaux traités. Ces effets comportementaux sont associés à une augmentation du nombre de neurones positifs à la cytochrome oxydase dans le noyau vestibulaire latéral du côté lésé, suggérant une implication du métabolisme énergétique neuronal par le traitement à l’hormone T4. |
| Dans le texte | |
|
Figure 4 Modèle proposé de l’interaction astrocyte-neurone et du transport de la T3 dans le cerveau. Tout d’abord l’hormone T4 (et à moindre mesure l’hormone T3) traverse la barrière hémato-encéphalique (BHE) grâce aux transporteurs OATP1 (organic anion transporting polypeptide 1) et MCT8 (monocarboxylate transporter 8). Trois scénarios sont possibles pour la suite : 1. L’hormone T4 est transportée dans les astrocytes par un transporteur encore inconnu. Dans les astrocytes, l’hormone T4 est convertie en T3 par l’enzyme D2. L’hormone T3 biologiquement active est ensuite transportée hors des astrocytes par un autre transporteur inconnu. 1b. L’hormone T3 depuis la BHE rejoint les neurones par MCT8. 2. L’assimilation neuronale de l’hormone T4 métabolisée en T3 par les astrocytes est facilitée par MCT8 puis l’hormone T3 exerce son action génomique en se liant à son récepteur nucléaire (TR). |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.