")
")
| Issue |
Med Sci (Paris)
Volume 42, Number 5, Mai 2026
|
|
|---|---|---|
| Page(s) | 431 - 435 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/2026075 | |
| Published online | 29 mai 2026 | |
La protéine de capside ORF2 du virus de l’hépatite E, clé de voûte de la persistance virale
The hepatitis E virus ORF2 capsid protein, a cornerstone of viral persistence
Université de Lille, CNRS, Inserm, CHU Lille, Institut Pasteur de Lille, U1019-UMR9017, Centre d’infection et d’immunité de Lille (CIIL), Lille, France
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Ces dernières années a émergé la théorie du « monde à ARN », selon laquelle les virus à ARN contemporains pourraient constituer des réminiscences de l’origine de la vie sur Terre. Des millions d’années d’évolution les ont conduits à devenir des parasites obligatoires, incapables de se multiplier en l’absence d’un hôte cellulaire. Cette longue évolution des virus à ARN, suivie d’une coévolution avec leurs hôtes, a conduit à une optimisation de leurs génomes, parfois longs de moins de 2 000 nucléotides et caractérisés par la présence de cadres ouverts de lecture (open reading frames, ORF) chevauchants. En conséquence, les protéines virales qu’ils codent sont elles aussi optimisées, fréquemment synthétisées sous forme de polyprotéines ou de protéines multifonctionnelles.
Le virus de l’hépatite E illustre cette stratégie évolutive. Ce virus est responsable de la plupart des cas d’hépatite aigüe dans le monde [1]. Il infecte plus de 20 millions de personnes chaque année et en tue 40 000 à 70 000. Il circule sous forme de huit génotypes, qui diffèrent par leur voie de transmission, leur distribution géographique et leur pathogénicité. En France, la séroprévalence est de 22 %, et le principal mode de contamination est la consommation de viande de porc ou de gibier pas ou peu cuite. L’infection par ce virus provoque généralement une hépatite aigüe. Celle-ci peut aussi évoluer vers une hépatite chronique chez les patients immunodéprimés infectés par le virus de génotype 3. En outre, l’infection des femmes enceintes par le virus de génotype 1 est mortelle dans 30 % des cas.
Le génome de ce virus est constitué d’un ARN simple brin de polarité positive, d’environ 7 200 nucléotides, qui code quatre protéines désignées par ORF1 à ORF4 [1, 15] (→).
(→) Voir m/s n° 12, 2018, page 1071
ORF1 est une polyprotéine de 1 693 acides aminés, dont le clivage en domaines fonctionnels reste débattu. Elle assure la réplication du génome viral grâce à ses activités ARN polymérase ARN-dépendante, hélicase, et méthyltransférase, mais elle contient également d’autres domaines dont les fonctions sont mal caractérisées [1]. ORF3 est une petite protéine de 113 acides aminés impliquée dans l’enveloppement et la sécrétion des particules virales. La protéine ORF4, encore peu étudiée, est spécifique du virus de génotype 1 et n’est exprimée que dans des conditions de stress du réticulum endoplasmique. Enfin, ORF2 est une protéine multifonctionnelle de 660 acides aminés qui remplit des rôles à la fois structurels et non structurels.
Sa fonction première est de constituer la capside virale assurant la protection physique de l’ARN viral [1]. Les résultats de nombreux travaux de recherche ont permis de lui attribuer également un rôle dans la modulation de la réponse antivirale des cellules infectées [2]. C’est dans ce contexte qu’une équipe de chercheurs a récemment publié un article visant à caractériser les mécanismes moléculaires de cette activité immunomodulatrice de ORF2 [3].
Les chercheurs ont d’abord utilisé un modèle d’expression ectopique de ORF2 (issue des génotypes 1 et 3) dans des cellules A549, dérivées d’un adénocarcinome de l’épithélium alvéolaire pulmonaire. Bien que ces cellules soient très différentes des hépatocytes, principales cellules cibles du virus, elles présentaient l’avantage d’être déficientes pour plusieurs récepteurs de reconnaissance des motifs (pattern recognition receptors, PRR), qui ont ainsi pu être réintroduits individuellement. Dans ce système, les auteurs ont montré que l’expression de la protéine ORF2 interfère avec l’induction des interférons de type I et III ainsi qu’avec l’expression des gènes stimulés par les interférons (interferon-stimulated genes, ISG), en inhibant certains composants des voies de signalisation en aval de PRR tels que MDA5 (melanoma differentiation-associated protein 5), RIG-I (retinoic acid-inducible gene I) et TLR3 (toll-like receptor 3), sans altérer la signalisation JAK (Janus kinase) / STAT (signal transducer and activator of transcription) induite par des interférons exogènes. Cette inhibition repose en partie sur une interaction directe entre la protéine ORF2 et la kinase adaptatrice TBK1 (TANK-binding kinase 1), interaction qui limite l’activation des facteurs de transcription IRF3 (interferon regulatory factor 3) et NF-κB (nuclear factor kB), entraînant une diminution de la production d’interférons et de cytokines pro-inflammatoires. Ces résultats sont cohérents avec ceux d’études précédentes montrant que ORF2 inhiberait la voie de signalisation impliquant NF-κB en interagissant avec β-TRCP (β-transducin repeat containing E3 ubiquitin protein ligase) [4], ou avec TMEM134 (transmembrane protein 134) [5]. Une autre étude a mis en évidence une interaction de ORF2 avec le complexe MAVS (mitochondrial antiviral signaling)-TBK1-IRF3 impliquant un motif riche en arginine (arginine-rich motif, ARM) situé à l’extrémité N-terminale de la protéine ORF2, et conduisant à une inhibition de la phosphorylation et de la translocation nucléaire de IRF3 [6]. Les expériences de co-immunoprécipitation ont permis de confirmer l’interaction entre ORF2 et TBK1 ainsi que l’importance du motif ARM, bien qu’une modélisation de cette interaction par AlphaFold2 n’ait pas permis d’identifier le rôle structural de ce motif [3]. L’absence d’analyses fonctionnelles utilisant une ORF2 porteuse d’une mutation du motif ARM constitue une limitation de l’étude.
Les chercheurs ont ensuite produit des génomes du virus déficients pour l’expression de ORF2 (ΔORF2) ou de ORF3 (ΔORF3), et montré que l’absence de la protéine ORF2 entraîne une induction accrue et prolongée de la réponse antivirale, associée à une diminution progressive de la réplication virale. Ces constats ont été faits aussi bien après électroporation des génomes viraux dans la lignée cellulaire hépatique HepG2/C3A1, que lors d’infections de cellules HepG2/C3A ou de cellules souches différenciées en hépatocytes avec des particules virales produites par trans-complémentation. Notons que la restriction de la réplication du virus mutant ΔORF2 a pu être partiellement levée par l’inhibition pharmacologique de TBK1, ce qui indique que l’hyperactivation de la réponse antivirale constitue un facteur limitant majeur de la progression de l’infection en l’absence de ORF2. Par ailleurs, les chercheurs ont montré la sensibilité accrue du mutant ΔORF2 aux traitements par l’interféron α et l’interféron λ dans la lignée hépatique Huh-7.52, déficiente pour la détection des ARN viraux via RIG-I. Ce constat suggère que ORF2 confère également une protection directe contre les effecteurs antiviraux induits par les ISG, indépendamment de son interaction avec TBK1. Ainsi, l’équilibre entre l’activation de la réponse cellulaire antivirale et son inhibition partielle par ORF2 apparaît comme une condition préalable à l’établissement d’une réplication persistante du virus.
L’ensemble des fonctions attribuées à ORF2 indique un rôle central de cette protéine dans la persistance virale. Trois mécanismes principaux peuvent être distingués. Le premier découle du rôle de ORF2 dans la formation de la capside du virus, qui protège physiquement l’ARN viral de son environnement, notamment de sa détection et de sa dégradation par les facteurs cellulaires antiviraux. Le second est la capacité de ORF2 à moduler la réponse antivirale induite par l’infection, soit par interaction avec TBK1 et d’autres facteurs de signalisation, soit par un mécanisme encore mal compris impliquant la localisation nucléaire de ORF2 [7, 8]. Enfin, le troisième repose sur le rôle de ORF2 comme leurre immunologique. ORF2 est en effet produite sous plusieurs formes : la forme infectieuse associée aux particules virales (ORF2i), ainsi que des formes glycosylées et clivées (ORF2g et ORF2c), abondamment sécrétées mais non associées aux virions [9, 10]. Ces formes sécrétées constituent la majorité des antigènes viraux détectés dans le sérum des patients infectés, et pourraient détourner la réponse humorale de la cible infectieuse. Dans la continuité des travaux de ces chercheurs, il serait pertinent de déterminer quelle(s) forme(s) de ORF2 antagonisent la réponse antivirale. Au-delà de ces trois mécanismes, ORF2 remplit d’autres fonctions dans le cycle viral. Exposée à la surface des particules virales non enveloppées, elle participe au tropisme viral. De concert avec ORF3, elle contribue également à la formation d’usines virales à proximité des endosomes de recyclage [11, 12], permettant la concentration des composants nécessaires à l’assemblage des néovirions. ORF2 apparaît ainsi comme une protéine multifonctionnelle, dont les rôles structurels et non structurels contribuent à la persistance du virus (Figure 1).
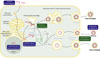 |
Figure 1 Les différents rôles de la protéine ORF2 au cours de l’infection par le virus de l’hépatite E. La protéine de capside ORF2 est synthétisée par traduction de l’ARN viral de 2,2 kb. Cette protéine existe sous trois formes : ORF2i (cytoplasmique ou nucléaire, impliquée notamment dans la formation de la nucléocapside), ORF2g et ORF2c (glycosylées, clivées et sécrétées). Les cadres verts indiquent les fonctions de ORF2 directement impliquées dans le cycle viral. Les cadres bleus indiquent ses rôles dans l’échappement immunitaire du virus. |
Les chercheurs ont ensuite réalisé une analyse transcriptomique comparative, à l’échelle unicellulaire (single cell RNA sequencing, scRNAseq), de cellules infectées par le virus de génotype 3 produisant ou non ORF2 et de cellules non infectées. Les auteurs ont fait le choix de réaliser cette analyse 56 heures après l’infection, alors qu’ils avaient précédemment montré une différence d’activation de la réponse antivirale entre le virus ΔORF2 et le virus non muté 5 jours après l’infection. Néanmoins, ce choix permet de mettre en lumière les évènements précurseurs ou précoces de cette réponse antivirale et de s’affranchir de ses effets sur la réplication du virus. La surexpression précoce de gènes associés à la réponse antivirale mise en évidence par cette analyse était plus marquée en l’absence de ORF2, comme l’était l’augmentation de l’activité métabolique, caractérisée par une activation accrue de la glycolyse et de la phosphorylation oxydative. Ces résultats sont cohérents avec des données cliniques et expérimentales obtenues précédemment chez l’Homme et chez le singe macaque [13, 14].
Pour terminer, les auteurs ont cherché à caractériser la réponse globale d’une population cellulaire à l’infection par le virus. À partir des données de séquençage transcriptomique, ils ont classé chaque cellule en fonction du degré d’expression d’un ensemble de 400 ISG. Cette analyse a permis d’identifier deux grands clusters cellulaires : d’une part, des cellules « actives », caractérisées par une forte expression des ISG et donc engagées dans une réponse antivirale, et d’autre part des cellules « non actives ». Sur cette base, quatre populations distinctes ont été définies : 1) des cellules non infectées, ne contenant pas d’ARN viral et non actives ; 2) des cellules infectées répondeuses, contenant l’ARN viral, actives et présentant une activité métabolique élevée ; 3) des cellules infectées non répondeuses, contenant l’ARN viral mais non actives ; et 4) des cellules non infectées mais actives. Cette dernière population pourrait correspondre soit à des cellules précédemment infectées ayant éliminé le virus grâce à leur réponse antivirale, hypothèse toutefois peu probable compte tenu de la précocité de l’analyse après l’infection, soit plus vraisemblablement à des cellules voisines de cellules infectées, dites « bystanders ». Par la détection de signaux inflammatoires paracrines émis par les cellules infectées, ces cellules bystanders acquièrent un phénotype pro-inflammatoire sans être elles-mêmes infectées. Ce mécanisme présente un double avantage pour l’hôte : à l’échelle individuelle, il permet à la cellule bystander de déclencher une réponse antivirale préventive, augmentant ses chances de contrôler efficacement une éventuelle infection ultérieure ; à l’échelle collective, il contribue à l’amplification du signal d’alerte, favorisant le recrutement et l’activation des cellules immunitaires au sein du tissu infecté (Figure 2). En présence du virus ΔORF2, la même analyse a révélé une activation accrue des ISG et une diminution du nombre de cellules contenant l’ARN viral, confirmant les conclusions établies précédemment. Les cellules bystanders des cellules infectées par le virus ΔORF2 ou par le virus non muté présentaient un degré d’activation des ISG comparable à celui des cellules infectées. Dans un contexte physiopathologique, cette stimulation paracrine revêt une importance particulière. Le foie est en effet composé d’environ deux tiers d’hépatocytes et d’un tiers de cellules non parenchymateuses incluant de nombreuses cellules immunitaires résidentes, dont plus de la moitié sont des macrophages d’origine embryonnaire (cellules de Kupffer) capables d’autorenouvellement. Lors d’une infection par le virus de l’hépatite E, ces cellules sont susceptibles de détecter les signaux d’alerte émis par les hépatocytes infectés et d’agir comme des relais puissants de l’inflammation, grâce à leur forte capacité de sécrétion de cytokines, tant en quantité qu’en diversité (Figure 2). En contrepartie, la production de ORF2 par la cellule infectée limiterait ce processus, favorisant la persistance virale et, potentiellement, l’établissement d’une infection chronique. Cet équilibre subtil entre activation de la réponse antivirale de l’hôte et stratégies virales d’échappement immunitaire illustre la coévolution des virus avec leur hôte. Bien qu’aucun traitement spécifique de l’infection par le virus de l’hépatite E ne soit actuellement recommandé, faire basculer cet équilibre en faveur de la réponse antivirale (par stimulation de l’immunité, inhibition directe du virus, ou par une combinaison des deux) constitue un objectif central des stratégies thérapeutiques en cours de développement.
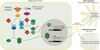 |
Figure 2 Inhibition de la réponse antivirale par la protéine ORF2 dans les hépatocytes infectés par le virus de l’hépatite E. La réplication du génome du virus par la polymérase virale ORF1 conduit à la formation d’intermédiaires ARN double brin. Ceux-ci, exposés dans le cytoplasme, peuvent être détectés par certains PRR (pattern recognition receptors) cellulaires (RIG-I, MDA5 ou TLR3), ce qui déclenche une cascade de signalisation aboutissant à la production de facteurs antiviraux et de cytokines pro-inflammatoires. Ces dernières ont pour rôle de transmettre un signal d’alerte paracrine aux hépatocytes adjacents non infectés (bystanders) et aux cellules immunitaires présentes dans le foie, telles que les macrophages résidents (cellules de Kupffer) ou les cellules dendritiques. La protéine virale ORF2, en se fixant notamment à TBK1 et à β-TRCP, aurait la capacité d’inhiber respectivement la voie de signalisation de l’interféron de type I et celle du TNF-α via NF-κB. D’autres mécanismes de blocage de la réponse antivirale par ORF2 restent à découvrir. |
Liens d’intérêt
Les auteurs déclarent ne pas avoir de lien d’intérêt.
Références
- Corneillie L, Mézière L, Montpellier C, et al. Update on the molecular and cellular biology of hepatitis E virus and therapeutic opportunities. Antivir Res 2026 ; 247 : 106353. [Google Scholar]
- Devhare P, Madiyal M, Mukhopadhyay C, et al. Interplay between hepatitis E virus and host cell pattern recognition receptors. Int J Mol Sci 2021 ; 22 : 9259. [Google Scholar]
- Mehnert AK, Stegmaier S, Alvarez CR, et al. The hepatitis E virus capsid protein ORF2 counteracts cell-intrinsic antiviral responses to enable persistent replication in cell culture. PLoS Pathog 2025 ; 21 : e1013516. [Google Scholar]
- Surjit M, Varshney B, Lal SK. The ORF2 glycoprotein of hepatitis E virus inhibits cellular NF-kB activity by blocking ubiquitination mediated proteasomal degradation of IkBa in human hepatoma cells. BMC Biochem 2012 ; 13 : 7. [Google Scholar]
- Tian Y, Huang W, Yang J, et al. Systematic identification of hepatitis E virus ORF2 interactome reveals that TMEM134 engages in ORF2-mediated NF-kB pathway. Virus Res 2017 ; 228 : 102-8. [Google Scholar]
- Lin S, Yang Y, Nan Y, et al. The capsid protein of hepatitis E virus inhibits interferon induction via its N-terminal arginine-rich motif. Viruses 2019 ; 11 : 1050. [Google Scholar]
- Hervouet K, Ferrié M, Ankavay M, et al. An arginine-rich motif in the ORF2 capsid protein regulates the hepatitis E virus lifecycle and interactions with the host cell. PLoS Pathog 2022 ; 18 : e1010798. [Google Scholar]
- Joshi G, Décembre E, Brocard J, et al. Plasmacytoid dendritic cell sensing of hepatitis E virus is shaped by both viral and host factors. Life Sci Alliance 2025 ; 8 : e202503256. [Google Scholar]
- Montpellier C, Wychowski C, Sayed IM, et al. Hepatitis E virus lifecycle and identification of three forms of the ORF2 capsid protein. Gastroenterology 2018 ; 154 : 211-23.e8. [Google Scholar]
- Yin X, Ying D, Lhomme S, et al. Origin, antigenicity, and function of a secreted form of ORF2 in hepatitis E virus infection. Proc Natl Acad Sci USA 2018 ; 115 : 4773-8. [Google Scholar]
- Bentaleb C, Hervouet K, Montpellier C, et al. The endocytic recycling compartment serves as a viral factory for hepatitis E virus. Cell Mol Life Sci 2022 ; 79 : 615. [Google Scholar]
- Metzger K, Bentaleb C, Hervouet K, et al. Processing and subcellular localization of the hepatitis E virus replicase: Identification of candidate viral factories. Front Microbiol 2022 ; 13 : 828636. [Google Scholar]
- Choi YH, Zhang X, Srinivasamoorthy G, et al. Transcriptome analysis in rhesus macaques infected with hepatitis E virus genotype 1/3 infections and genotype 1 re-infection. PLoS One 2020 ; 15 : e0237618. [Google Scholar]
- Munshi SU, Taneja S, Bhavesh NS, et al. Metabonomic analysis of hepatitis E patients shows deregulated metabolic cycles and abnormalities in amino acid metabolism. J Viral Hepat 2011 ; 18 : e591-602. [Google Scholar]
- Ankavay M, Dubuisson J, Cocquerel L. Le virus de l’hépatite E : un virus méconnu qui se dévoile. Med Sci (Paris) 2018 ; 34 : 1071-8. [Google Scholar]
La lignée cellulaire HepG2 est une lignée de cellules cancéreuses du foie humain. C3A désigne un dérivé clonal de cette lignée.
Autre lignée cellulaire dérivée d’un hépatocarcinome humain.
© 2026 médecine/sciences – Inserm
 Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l’utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l’utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Liste des figures
|
Figure 1 Les différents rôles de la protéine ORF2 au cours de l’infection par le virus de l’hépatite E. La protéine de capside ORF2 est synthétisée par traduction de l’ARN viral de 2,2 kb. Cette protéine existe sous trois formes : ORF2i (cytoplasmique ou nucléaire, impliquée notamment dans la formation de la nucléocapside), ORF2g et ORF2c (glycosylées, clivées et sécrétées). Les cadres verts indiquent les fonctions de ORF2 directement impliquées dans le cycle viral. Les cadres bleus indiquent ses rôles dans l’échappement immunitaire du virus. |
| Dans le texte | |
|
Figure 2 Inhibition de la réponse antivirale par la protéine ORF2 dans les hépatocytes infectés par le virus de l’hépatite E. La réplication du génome du virus par la polymérase virale ORF1 conduit à la formation d’intermédiaires ARN double brin. Ceux-ci, exposés dans le cytoplasme, peuvent être détectés par certains PRR (pattern recognition receptors) cellulaires (RIG-I, MDA5 ou TLR3), ce qui déclenche une cascade de signalisation aboutissant à la production de facteurs antiviraux et de cytokines pro-inflammatoires. Ces dernières ont pour rôle de transmettre un signal d’alerte paracrine aux hépatocytes adjacents non infectés (bystanders) et aux cellules immunitaires présentes dans le foie, telles que les macrophages résidents (cellules de Kupffer) ou les cellules dendritiques. La protéine virale ORF2, en se fixant notamment à TBK1 et à β-TRCP, aurait la capacité d’inhiber respectivement la voie de signalisation de l’interféron de type I et celle du TNF-α via NF-κB. D’autres mécanismes de blocage de la réponse antivirale par ORF2 restent à découvrir. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.