")
")
| Issue |
Med Sci (Paris)
Volume 35, Number 10, Octobre 2019
|
|
|---|---|---|
| Page(s) | 746 - 748 | |
| Section | Le Magazine | |
| DOI | https://doi.org/10.1051/medsci/2019145 | |
| Published online | 18 October 2019 | |
Un nouveau talon d’Achille du bacille de la tuberculose
A new Achilles’ heel in the tuberculosis bacillus
Institut de pharmacologie et biologie structurale (IPBS), université de Toulouse, CNRS, UPS, 205 route de Narbonne, 31000 Toulouse, France
*
This email address is being protected from spambots. You need JavaScript enabled to view it.
La tuberculose, infection principalement pulmonaire due à la bactérie Mycobacterium tuberculosis, est la maladie infectieuse due à un agent unique la plus meurtrière à l’échelle mondiale1. L’expansion de souches de M. tuberculosis multi-résistantes, voire totalement résistantes, aux antibiotiques fait craindre l’apparition d’une pandémie incontrôlable. Ainsi, bien que la tuberculose soit, dans l’imaginaire collectif, une maladie associée au passé, elle reste un problème de santé majeur, et la recherche de nouvelles pistes thérapeutiques est plus que jamais nécessaire. Dans une étude récente, menée par un consortium associant notre équipe et deux équipes du Laboratoire européen de biologie moléculaire (EMBL) à Hambourg et de l’Institut Francis Crick à Londres, nous avons identifié une nouvelle piste thérapeutique, fondée sur l’activation d’un système toxine/antitoxine (TA) pouvant entraîner le « suicide » des cellules de M. tuberculosis [1].
Les systèmes toxine/antitoxine
Chez les bactéries, les systèmes TA sont des modules formés d’un gène codant une toxine protéique, capable d’intoxiquer la cellule productrice en bloquant un processus essentiel de son métabolisme (synthèse des protéines par exemple), et d’un élément d’immunité contre cette toxine, l’antitoxine, qui peut, en inhibant la production et/ou l’activité de la toxine, protéger la bactérie productrice [2, 3]. Ces systèmes sont regroupés en diverses familles (de I à VI) selon la nature de l’antitoxine (protéine ou ARN), et le mécanisme d’inactivation de la toxine. Par exemple, dans les systèmes TA de type II, les plus étudiés, l’antitoxine est une protéine qui se lie directement à la toxine pour l’inactiver. Pour les systèmes TA de type IV, la toxine est aussi une protéine, mais elle empêche l’action de la toxine sans interaction directe avec celle-ci. La clé du fonctionnement des systèmes TA est que la toxine et l’antitoxine sont stables en conditions physiologiques standard, mais que l’antitoxine est déstabilisée dans certaines conditions de stress, libérant l’activité de la toxine qui bloque alors la croissance ou peut même entraîner la mort des bactéries soumises au stress. Initialement identifiés comme des systèmes « d’addiction » à des plasmides, les systèmes TA sont présents dans la plupart des génomes bactériens, et ils participent à diverses fonctions biologiques, dont la stabilisation de réplicons ou d’îlots génomiques, la lutte contre les infections par les bactériophages, ou l’entrée en persistance des bactéries. Cependant, le rôle effectif de la plupart des systèmes TA reste une question ouverte [4, 5]. Ceci est particulièrement vrai pour M. tuberculosis, dont le génome est très riche en systèmes TA. En effet, on retrouve dans le génome de M. tuberculosis plus de 80 systèmes TA, principalement de type II, dont un grand nombre sont portés par des îlots génomiques acquis par transfert horizontal chez l’ancêtre de M. tuberculosis [6].
Le système MbcTA de M. tuberculosis
Utiliser les toxines des systèmes TA comme outils antibactériens a été souvent proposé (voir par exemple [7, 8]). Cependant, la découverte du lien entre certains systèmes TA et la persistance a remis cette idée en question, puisque l’activation d’une toxine risquerait de rendre les cellules bactériennes tolérantes aux antibiotiques. Afin d’éviter cet écueil, nous avons utilisé les résultats d’études de mutagénèse à saturation de M. tuberculosis, qui ont montré que parmi les nombreux systèmes TA présents chez cette espèce, seulement trois possèdent un gène d’antitoxine qui est essentiel [9], c’est-à-dire qui ne peut pas être inactivé génétiquement. Nous avons fait l’hypothèse que cela était dû à une toxicité létale de la toxine associée, et nous avons analysé plus avant l’un de ces systèmes, codé par l’opéron formé des gènes appelés Rv1990c et Rv1989c. Nous avons alors confirmé que l’opéron Rv1990c-Rv1989c code bien un système TA, mais surtout que l’action de la toxine a un effet bactéricide chez M. tuberculosis (Figure 1). Nous avons alors nommé ce système MbcTA (mycobacterial cidal toxin/antitoxin).
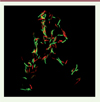 |
Figure 1. MbcT est un système TA bactéricide chez M. tuberculosis. Les cellules d’une souche de M. tuberculosis délétée de l’opéron MbcTA (MtbDTA), transformée par un plasmide exprimant la toxine Rv1989c sous contrôle d’un promoteur inductible par la tétracycline, ont été traitées au temps 0 par addition d’anhydro-tétracycline (ATc). Après quatre jours d’induction de la production de toxine, les cellules sont marquées avec le kit LIVE/DEAD BacLight (Syto 9 ; iodure de propidium, PI) colorant en vert les cellules viables et en rouge les cellules dont la membrane est perméabilisée (non-viables). Les cellules sont observées en microscopie à fluorescence. |
MbcT est une NAD+ phosphorylase
Afin d’analyser le mécanisme d’action de cette toxine, nous avons déterminé sa structure tridimensionnelle. Il a été possible de surproduire et cristalliser un complexe de protéines MbcTA, démontrant qu’il s’agit d’un système de type II. Ce complexe a une organisation en dodécamère (trimère de tétramères [MbcTA]2). La signification biologique de cette association reste à comprendre. En revanche, bien que la séquence primaire de la toxine MbcT ne présente aucune similarité avec des protéines de fonction connue, sa structure tridimensionnelle ressemble à celle des exotoxines bactériennes de la famille des ADP-ribosyl-transférases, comme les toxines diphtérique ou cholérique. L’analyse biochimique et métabolomique de MbcT a montré que cette toxine a une activité originale. En effet, elle catalyse la dégradation du coenzyme essentiel NAD+ par phosphorolyse, produisant du nicotinamide et de l’ADP-ribose-1’’-phosphate (Figure 2). Une telle activité de phosphorolyse du NAD+ n’avait encore jamais été identifiée. En accord avec cette activité, nous avons montré que l’induction de MbcT conduit à un effondrement du contenu en NAD+ dans les cellules de M. tuberculosis.
 |
Figure 2. MbcT est une NAD+ phosphorylase. Mécanisme de phosphorolyse du NAD+, catalysée par la toxine MbcT. Appr1p : ADP-ribose-1’’-phosphate ; NAA : nicotinamide. |
L’activation de MbcTA peut protéger contre l’infection par M. tuberculosis
L’étape suivante de notre travail a été d’apporter « la preuve de concept » que l’activation de MbcT pourrait être utilisée dans une stratégie thérapeutique, en complément de l’antibiothérapie antituberculeuse. Nous avons utilisé une souche de M. tuberculosis dépourvue (par génie génétique) du système MbcTA chromosomique et exprimant une copie de MbcT sous le contrôle d’un promoteur inductible par des analogues de la tétracycline, l’anhydro-tétracycline (ATc) et la doxycycline. Nous avons alors montré que l’addition d’ATc à des macrophages humains en culture infectés par cette souche permet de réduire la charge bactérienne, démontrant que l’action bactéricide est conservée dans la niche cellulaire principale de M. tuberculosis que sont les macrophages de l’hôte. De plus, l’administration orale de doxycycline à des souris infectées par cette souche diminue la charge bactérienne dans les poumons, et cet effet est synergique avec celui d’un traitement par l’isoniazide, un antibiotique de première ligne utilisé dans le traitement de la tuberculose.
L’ensemble de ces travaux ouvre la voie à l’analyse d’une nouvelle classe d’enzymes, les NAD+ phosphorylases, présentes non seulement chez M. tuberculosis, mais aussi chez d’autres espèces bactériennes [10]. De plus, nos travaux en cours visent à exploiter le système MbcT pour identifier des molécules capables d’activer la toxine, soit en interférant avec le complexe MbcTA, soit en stimulant la dégradation de l’antitoxine MbcA. Pour cela, nous développons des cribles sur cellules entières, dont nous espérons qu’ils identifieront de nouvelles molécules antituberculeuses.
Liens d’intérêt
Les auteurs déclarent n’avoir aucun lien d’intérêt concernant les données publiées dans cet article.
Rapport de l’Organisation mondiale de la santé 2018.
Références
- Freire DM, Gutierrez C, Garza-Garcia A, et al. An NAD+ phosphorylase toxin triggers Mycobacterium tuberculosis cell death. Mol Cell 2019 ;73(1282–91) :e8. [Google Scholar]
- Harms A, Brodersen DE, Mitarai N, Gerdes K. Toxins, targets, and triggers: an overview of toxin-antitoxin biology. Mol Cell 2018 ;70 :768–784. [CrossRef] [PubMed] [Google Scholar]
- Page R, Peti W. Toxin-antitoxin systems in bacterial growth arrest and persistence. Nat Chem Biol 2016 ;12 :208–214. [Google Scholar]
- Magnuson RD. Hypothetical functions of toxin-antitoxin systems. J Bacteriol 2007 ;189 :6089–6092. [CrossRef] [PubMed] [Google Scholar]
- Van Melderen L. Toxin-antitoxin systems: why so many, what for? Curr Opin Microbiol 2010 ;13 :781–785. [CrossRef] [PubMed] [Google Scholar]
- Sala A, Bordes P, Genevaux P. Multiple toxin-antitoxin systems in Mycobacterium tuberculosis. Toxins (Basel) 2014 ;6 :1002–1020. [CrossRef] [PubMed] [Google Scholar]
- Chan WT, Balsa D, Espinosa M. One cannot rule them all: Are bacterial toxins-antitoxins druggable? FEMS Microbiol Rev 2015 ;39 :522–540. [CrossRef] [PubMed] [Google Scholar]
- Lee KY, Lee BJ. Structure, biology, and therapeutic application of toxin-antitoxin systems in pathogenic bacteria. Toxins (Basel) 2016 ;8 :E305. [CrossRef] [PubMed] [Google Scholar]
- DeJesus MA, Gerrick ER, Xu W, et al. Comprehensive essentiality analysis of the Mycobacterium tuberculosis genome via saturating transposon mutagenesis. MBio 2017 ;8 :e02133–16. [CrossRef] [PubMed] [Google Scholar]
- Skjerning RB, Senissar M, Winter KS, et al. The RES domain toxins of RES-Xre toxin-antitoxin modules induce cell stasis by degrading NAD+. Mol Microbiol 2019 ;111 :221–236. [CrossRef] [PubMed] [Google Scholar]
© 2019 médecine/sciences – Inserm
 Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (http://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l'utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (http://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l'utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Liste des figures
|
Figure 1. MbcT est un système TA bactéricide chez M. tuberculosis. Les cellules d’une souche de M. tuberculosis délétée de l’opéron MbcTA (MtbDTA), transformée par un plasmide exprimant la toxine Rv1989c sous contrôle d’un promoteur inductible par la tétracycline, ont été traitées au temps 0 par addition d’anhydro-tétracycline (ATc). Après quatre jours d’induction de la production de toxine, les cellules sont marquées avec le kit LIVE/DEAD BacLight (Syto 9 ; iodure de propidium, PI) colorant en vert les cellules viables et en rouge les cellules dont la membrane est perméabilisée (non-viables). Les cellules sont observées en microscopie à fluorescence. |
| Dans le texte | |
|
Figure 2. MbcT est une NAD+ phosphorylase. Mécanisme de phosphorolyse du NAD+, catalysée par la toxine MbcT. Appr1p : ADP-ribose-1’’-phosphate ; NAA : nicotinamide. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.