")
")
| Issue |
Med Sci (Paris)
Volume 30, Number 10, Octobre 2014
|
|
|---|---|---|
| Page(s) | 828 - 830 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/20143010003 | |
| Published online | 14 October 2014 | |
Les complexes de Merkel
De l’histologie ancienne aux mécanismes moléculaires du toucher
Merkel complexes: from old histology to molecular mechanisms of touch
Aix-Marseille université, CNRS, UMR 7286, Centre de recherche en neurobiologie-neurophysiologie de Marseille, Faculté de médecine, campus Nord 51, boulevard Pierre Dramard, 13344, Marseille Cedex 15, France
*
This email address is being protected from spambots. You need JavaScript enabled to view it.
Le complexe de Merkel cutané
Ouvrons un ouvrage d’histologie cutanée : les corpuscules de Pacini, Ruffini, Meissner et les complexes de Merkel décrits depuis la fin du xixe siècle forment les structures du toucher. Le toucher englobe plusieurs modalités sensorielles (la caresse, l’exploration des formes, des textures mais aussi la douleur) qui résultent toutes des propriétés de nos récepteurs. Chaque récepteur tactile se caractérise par le stimulus qui l’active (une pression ponctuelle, un étirement, un glissement, une vibration, une piqûre), par la taille du champ récepteur, et par la durée de la réponse lors d’une pression continue. Un récepteur à adaptation lente fonctionne aussi longtemps que la pression se maintient, un récepteur à adaptation rapide ne fonctionne que lors d’un changement de pression. À partir des années 1960, l’enregistrement d’une fibre sensorielle pendant une stimulation tactile a permis de préciser le seuil de sensibilité, la taille du champ récepteur et la vitesse d’adaptation de chaque récepteur. Ces travaux remarquables d’électrophysiologie sont à la base des hypothèses actuelles sur les mécanismes du toucher (voir pour revue [1]).
Néanmoins, on ne sait toujours pas quelles sont les molécules qui convertissent la déformation mécanique en signal biologique, quelle est la part respective des terminaisons nerveuses et des autres cellules des complexes, ni comment se fait la communication entre ces éléments. Ces questions ont été en partie résolues ces derniers mois par trois équipes travaillant sur la mécanosensibilité des complexes de Merkel de souris et sur les vibrisses de rat. Leurs résultats ont fait l’objet de publications dans les revues Nature et Cell.
Un complexe de Merkel est composé de quelques kératinocytes associés à des cellules de Merkel, elles-mêmes connectées aux terminaisons d’une fibre sensorielle Aβ. Les ramifications d’une fibre Aβ contactent ainsi 10 à 50 cellules de Merkel (Figure 1). Les cellules de Merkel sont issues de la lignée épidermique [2, 3], elles contiennent des neuropeptides et le transporteur du glutamate VGlut2. L’application d’une pression sur la peau déforme les cellules du complexe et déclenche dans la fibre Aβ une bouffée de potentiels d’action, puis une réponse à fréquence plus faible. Les complexes de Merkel sont donc des récepteurs mécaniques à bas seuil, au champ récepteur étroit et qui s’adaptent lentement à une pression (Figure 1). Ils vont transmettre des informations sur la forme et la texture des objets ; ils sont donc essentiels à la sensibilité tactile des mains chez l’homme et des coussinets plantaires chez la souris. Les cellules de Merkel sont indispensables à l’établissement de ces réponses et celles-ci tendent à disparaître dans un modèle de souris dépourvues de cellules de Merkel [4].
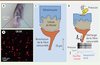 |
Figure 1. Le complexe tactile de Merkel. A. La peau glabre de la face ventrale d’une patte de souris, et en particulier les coussinets plantaires, contiennent des complexes de Merkel. B. Vue de dessus d’un fragment de peau étalée issu de la patte ventrale de souris. On visualise les cellules de Merkel (en rouge) par un marquage immunohistochimique dirigé contre la cytokératine 20 (CK20). C. Un complexe de Merkel est formé par l’association de trois types de cellules : des kératinocytes, des cellules de Merkel et la ramification d’une fibre nerveuse Aβ. Chaque fibre contacte par ses ramifications 10 à 50 cellules de Merkel. D. Une pression exercée sur la peau déforme les cellules du complexe de Merkel. L’enregistrement de l’activité d’une fibre Aβ pendant la déformation mécanique permet de mesurer le seuil de sensibilité du complexe (trait pointillé vert). Lors d’une variation de pression suivie d’une phase de pression maintenue, le complexe de Merkel répond d’abord avec une bouffée de potentiels d’action (trait pointillé rouge), puis avec une fréquence plus faible (trait pointillé bleu). La baisse de fréquence constitue l’adaptation au stimulus. |
Rôle du canal Piezo 2 des cellules de Merkel lors d’une réponse tactile
Les deux articles publiés dans la revue Nature [5, 6] portent sur les complexes de Merkel de la souris. Pour identifier les cellules de Merkel, les auteurs ont utilisé des souris qui expriment la GFP (green fluorescent protein) sous le contrôle du promoteur du gène Atoh1, qui code pour un facteur de transcription spécifique des cellules de Merkel [2, 3]. Il est ainsi possible d’isoler des cellules de Merkel et de tester leur réponse à une stimulation mécanique [7]. Il apparaît que les cellules de Merkel expriment des courants mécanosensibles qui résultent de l’activité du canal Piezo 2. Les canaux Piezo sont des canaux ioniques mécanosensibles excitateurs décrits très récemment chez les mammifères [8, 9].
Afin de réaliser leur étude, les chercheurs de l’équipe d’Ardem Patapoutian (the Scripps research Institute, États-Unis) ont généré plusieurs souches de souris génétiquement modifiées. Les souris chez lesquelles la GFP est fusionnée au canal Piezo2 confirment que ce dernier est exprimé à la fois dans les cellules de Merkel et dans la terminaison des fibres Aβ. Il est donc à priori difficile de distinguer le rôle respectif de ce canal dans chaque localisation. C’est pourquoi l’équipe a généré des souris dont le gène Piezo 2 a été inactivé spécifiquement dans les cellules épithéliales (souris Piezo2flox/flox;Krt14Cre, nommées souris Piezo 2CKO), ce qui permet de supprimer la transduction mécanique dans les cellules de Merkel. La réponse tactile a été étudiée ex vivo sur une préparation isolant la peau de la patte postérieure et les branches du nerf saphène. Deux modifications importantes sont observées chez ces souris : la fréquence de décharge associée à la première réponse en bouffée diminue légèrement et surtout, l’activité associée à la phase d’adaptation lente diminue considérablement (Figure 2). D’un point de vue comportemental, ces souris perdent en partie leur capacité de réaction à un toucher léger.
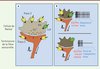 |
Figure 2. Modèle de transduction mécanique dans le complexe de Merkel. A. Une stimulation mécanique appliquée sur le complexe active les canaux ioniques cationiques mécanosensibles Piezo 2 des cellules de Merkel. L’entrée des cations dépolarise la cellule de Merkel et active des canaux calciques dépendants du potentiel (CaV). On suppose que cette augmentation de Ca2+ entraîne la libération d’un neuromédiateur (encore inconnu) qui active des récepteurs sur la terminaison de la fibre sensorielle Aβ. Des données immunohistochimiques suggèrent également la présence de canaux Piezo 2 sur la terminaison des fibres Aβ, mais l’enregistrement de courants mécanosensibles à ce niveau n’a pas été réalisé. Le rôle des kératinocytes (non représentés) reste inconnu. B. Schéma du haut : en conditions normales, la réponse du complexe entraîne au niveau de la fibre sensorielle Aβ une décharge à bas seuil d’activation, avec une bouffée initiale de potentiels d’action et une adaptation lente à la pression. Schéma du bas : en absence de canaux Piezo 2 dans la cellule de Merkel, la fréquence de la bouffée initiale est réduite et on perd la phase d’adaptation lente. Le complexe perd donc sa capacité à coder pour une pression continue. |
L’équipe d’Ellen Lumpkin (Columbia University, États-Unis) a adopté une stratégie d’optogénétique pour montrer la contribution des cellules de Merkel à une réponse tactile. Un canal cationique (channel rhodopsin 2, activateur), activé par une lumière bleue, a été introduit par un vecteur spécifique dans les cellules de Merkel. Son activation induit des potentiels d’action sur la fibre sensorielle Aβ, démontrant pour la première fois que la stimulation d’une cellule de Merkel est suffisante pour induire un message sensoriel tactile. Une pompe à proton (Archaerhodopsin 3, inhibiteur), activée par une lumière verte, a également été utilisée. L’inhibition des cellules de Merkel pendant une stimulation mécanique diminue considérablement la réponse des fibres tactiles Aβ. Ces résultats montrent que les cellules de Merkel contribuent à la réponse de la fibre sensorielle. Les canaux mécanosensibles des fibres Aβ ne sont donc pas suffisants pour produire une réponse normale. Pour assurer une dépolarisation prolongée, les cellules de Merkel mettent en jeu des canaux calciques dépendant du voltage qui sont activés par l’ouverture des canaux Piezo 2. C’est ce mécanisme de coopération entre canaux ioniques qui assure une réponse prolongée et donc l’adaptation lente du complexe au stimulus mécanique (Figure 2).
Information tactile des vibrisses et cellules de Merkel
La dernière étude, publiée dans Cell par l’équipe de Jianguo Gu (University of Cincinnati, États-Unis), porte sur la transduction mécanique dans les follicules des moustaches du rat [10]. Il s’agit d’un système qui n’a pas d’équivalent chez l’homme, mais qui a souvent servi de modèle pour caractériser le codage du message sensoriel tactile et son traitement cortical [11]. À l’intérieur du follicule, un anneau de cellules de Merkel est associé à des branches de fibres nerveuses de type Aβ. Les auteurs sont parvenus à enregistrer l’activité des cellules de Merkel lors d’une déformation mécanique du follicule et à montrer l’existence de courants mécanosensibles également portés par le canal Piezo 2. Ils ont montré que l’activité de ces canaux est nécessaire pour déclencher une réponse sensorielle lors du déplacement d’une moustache. Ainsi, dans ce système, le codage de l’information tactile semble aussi dépendre de la mécanosensibilité des cellules de Merkel. Néanmoins, certaines questions subsistent, par exemple, pourquoi l’élimination des cellules de Merkel entraîne-t-elle une réduction de la sensibilité tactile des pattes mais pas de l’efficacité de l’exploration spatiale par les vibrisses [12] ?
En conclusion, deux avancées conceptuelles majeures sont à retenir de ces travaux : (1) le codage de l’information tactile cutanée transmise par les complexes de Merkel nécessite le fonctionnement des cellules de Merkel qui confèrent les propriétés d’adaptation lente de la réponse ; et (2) au sein de ces cellules de Merkel, Piezo 2 est le transducteur mécanique. L’altération pathologique de ces cellules devrait donc entraîner un déficit tactile, en particulier lors de l’exploration haptique des objets qui s’effectue sous de faibles pressions. Il reste à comprendre quels médiateurs assurent la communication entre les cellules du complexe de Merkel, et à engager des études comparables dans les autres corpuscules du toucher.
Liens d’intérêt
Les auteurs déclarent n’avoir aucun lien d’intérêt concernant les données publiées dans cet article.
Remerciements
Les auteurs remercient Bertrand Coste et Patrick Delmas pour leurs commentaires et leurs suggestions.
Références
- Johansson RS, Vallbo AB. Tactile sensory coding in the glabrous skin of the human hand. Trends Neurosci 1983 ; 6 : 27–32. [CrossRef] [Google Scholar]
- Van Keymeulen A, Mascre G, Youseff KK, et al. Epidermal progenitors give rise to Merkel cells during embryonic development and adult homeostasis. J Cell Biol 2009 ; 187 : 91–100. [CrossRef] [PubMed] [Google Scholar]
- Morrison KM, Miesegaes GR, Lumpkin EA, Maricich SM. Mammalian Merkel cells are descended from the epidermal lineage. Dev Biol 2009 ; 336 : 76–83. [CrossRef] [PubMed] [Google Scholar]
- Maricich SM, Wellnitz SA, Nelson AM, et al. Merkel cells are essential for light-touch responses. Science 2009 ; 324 : 1580–1582. [CrossRef] [PubMed] [Google Scholar]
- Woo SH, Ranade S, Weyer AD, et al. Piezo2 is required for Merkel-cell mechanotransduction. Nature 2014 ; 509 : 622–626. [CrossRef] [PubMed] [Google Scholar]
- Maksimovic S, Nakatani M, Baba Y, et al. Epidermal Merkel cells are mechanosensory cells that tune mammalian touch receptors. Nature 2014 ; 509 : 617–621. [CrossRef] [PubMed] [Google Scholar]
- Hao J, Delmas P. Recording of mechanosensitive currents using piezoelectrically driven mechanostimulator. Nat Protoc 2011 ; 6 : 979–990. [CrossRef] [PubMed] [Google Scholar]
- Coste B. Détecter la pression ? Identification de deux protéines activées par les forces mécaniques. Med Sci (Paris) 2011 ; 27 : 17–19. [CrossRef] [EDP Sciences] [PubMed] [Google Scholar]
- Coste B. Les protéines Piezo forment une nouvelle famille de canaux ioniques activés mécaniquement. Med Sci (Paris) 2012 ; 28 : 1056–1057. [CrossRef] [EDP Sciences] [PubMed] [Google Scholar]
- Ikeda R, Cha M, Ling J, et al. Merkel cells transduce and encode tactile stimuli to drive aβ-afferent impulses. Cell 2014 ; 157 : 664–675. [CrossRef] [PubMed] [Google Scholar]
- Estebanez L, El Boustani S, Destexhe A, Shulz DE. Ce que les vibrisses disent au cerveau tactile. Med Sci (Paris) 2014 ; 30 : 93–98. [CrossRef] [EDP Sciences] [PubMed] [Google Scholar]
- Maricich SM, Morrison KM, Mathes EL, Brewer BM. Rodents rely on Merkel cells for texture discrimination tasks. J Neurosci 2012 ; 32 : 3296–3300. [CrossRef] [PubMed] [Google Scholar]
© 2014 médecine/sciences – Inserm
Liste des figures
|
Figure 1. Le complexe tactile de Merkel. A. La peau glabre de la face ventrale d’une patte de souris, et en particulier les coussinets plantaires, contiennent des complexes de Merkel. B. Vue de dessus d’un fragment de peau étalée issu de la patte ventrale de souris. On visualise les cellules de Merkel (en rouge) par un marquage immunohistochimique dirigé contre la cytokératine 20 (CK20). C. Un complexe de Merkel est formé par l’association de trois types de cellules : des kératinocytes, des cellules de Merkel et la ramification d’une fibre nerveuse Aβ. Chaque fibre contacte par ses ramifications 10 à 50 cellules de Merkel. D. Une pression exercée sur la peau déforme les cellules du complexe de Merkel. L’enregistrement de l’activité d’une fibre Aβ pendant la déformation mécanique permet de mesurer le seuil de sensibilité du complexe (trait pointillé vert). Lors d’une variation de pression suivie d’une phase de pression maintenue, le complexe de Merkel répond d’abord avec une bouffée de potentiels d’action (trait pointillé rouge), puis avec une fréquence plus faible (trait pointillé bleu). La baisse de fréquence constitue l’adaptation au stimulus. |
| Dans le texte | |
|
Figure 2. Modèle de transduction mécanique dans le complexe de Merkel. A. Une stimulation mécanique appliquée sur le complexe active les canaux ioniques cationiques mécanosensibles Piezo 2 des cellules de Merkel. L’entrée des cations dépolarise la cellule de Merkel et active des canaux calciques dépendants du potentiel (CaV). On suppose que cette augmentation de Ca2+ entraîne la libération d’un neuromédiateur (encore inconnu) qui active des récepteurs sur la terminaison de la fibre sensorielle Aβ. Des données immunohistochimiques suggèrent également la présence de canaux Piezo 2 sur la terminaison des fibres Aβ, mais l’enregistrement de courants mécanosensibles à ce niveau n’a pas été réalisé. Le rôle des kératinocytes (non représentés) reste inconnu. B. Schéma du haut : en conditions normales, la réponse du complexe entraîne au niveau de la fibre sensorielle Aβ une décharge à bas seuil d’activation, avec une bouffée initiale de potentiels d’action et une adaptation lente à la pression. Schéma du bas : en absence de canaux Piezo 2 dans la cellule de Merkel, la fréquence de la bouffée initiale est réduite et on perd la phase d’adaptation lente. Le complexe perd donc sa capacité à coder pour une pression continue. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.