")
")
| Issue |
Med Sci (Paris)
Volume 27, Number 1, Janvier 2011
|
|
|---|---|---|
| Page(s) | 31 - 33 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/201127131 | |
| Published online | 10 February 2011 | |
Les microARN, une nouvelle voie de signalisation cellulaire empruntée par le resvératrol
Resveratrol acts by modulating miRNAs
Université de Bourgogne, Inserm UMR 866, 6, boulevard Gabriel, 21000 Dijon, France
*
This email address is being protected from spambots. You need JavaScript enabled to view it.
Les microARN (miARN), découverts en 1993 dans le laboratoire de V. Ambros [1], ont d’abord été identifiés comme régulateurs du développement chez Caenorhabditis elegans. Les recherches récentes confirment que ces petits ARN non codants simple brin sont des éléments de signalisation cellulaire fondamentaux dans la régulation de processus tels que le développement, la différenciation ou la prolifération cellulaires. Ces ARN de 22 nucléotides en moyenne s’apparient de façon spécifique à des ARN messagers cibles entraînant le blocage de leur traduction (en cas de complémentarité parfaite) ou la dégradation des transcrits (s’il existe un ou quelques mésappariement[s]). Ils pourraient aussi agir au niveau transcriptionnel pour certains gènes. La mutation de gènes codant certains miARN entraîne des processus oncogéniques [2]. Cependant, les connaissances de la régulation des gènes codant ces miARN sont encore limitées.
Propriétés anti-inflammatoires et antitumorales du resvératrol
Notre équipe étudie les propriétés anti-inflammatoires et anticancéreuses du resvératrol, un phytophénol bien connu comme microconstituant alimentaire d’origine végétale (raisin, vin rouge, graines d’arachide, myrtille, etc.) et dont on connaît les propriétés protectrices vis-à-vis de pathologies majeures telles que l’athérosclérose, le cancer, les neurodégénérescences, ou encore l’inflammation. Selon le type de maladie, le resvératrol agirait principalement et respectivement en tant qu’anti-oxydant, inhibiteur de cyclines (associé ou non à un processus d’apoptose), activateur de sirtuines [3] ou encore inhibiteur de cyclo-oxygénases [4]. La signalisation induite par le resvératrol peut impliquer différents récepteurs, dont des intégrines, des récepteurs activateurs des MAP (mitogen-activated protein) kinases, les récepteurs de mort, les facteurs de transcription NFκB et NFR2 (NFE2 related factor 2), etc. La possible implication des miARN dans les mécanismes protecteurs exercés par le resvératrol n’avait pas été explorée.
Effets du resvératrol sur l’expression de mi ARN impliqués dans la tumorigenèse
Dans deux articles récents, nous venons de montrer, en collaboration avec le laboratoire de C. Croce (Columbus, Ohio, États-Unis), que le resvératrol module l’expression de miARN impliqués dans la tumorigenèse. Le premier travail montre que dans la lignée cellulaire THP-1 dérivée de monocytes sanguins humains, le resvératrol augmente l’expression du miARN 663 (miR-663) [5]. Ce dernier cible les transcrits de JunB et Jun D, et donc par voie de conséquence diminue l’activité du facteur de transcription AP-1 (activating protein -1), complexe constitué de Jun B et Jun D. En accord avec les propriétés anti-inflammatoires du resvératrol [6], la surexpression de miR-663 diminue l’expression de gènes impliqués dans la production de médiateurs pro-inflammatoires tels que cytokines, prostaglandines, espèces réactives de l’oxygène, etc. De plus, le resvératrol diminue le niveau d’expression d’un miARN pro-oncogénique et pro-inflammatoire, le miR-155. Ce microARN est par ailleurs activé par le LPS (lipopolysaccharide de paroi bactérienne, un puissant agent pro-inflammatoire) [7]. Cette régulation négative de miR-155 est la conséquence de la surexpression de miR-663 comme le montre la transfection des cellules THP-1 avec le miR-663 qui entraîne la baisse du niveau de miR-155. Aucune diminution d’expression de miR-155 n’est obtenue avec un miR-663 muté (Figure 1). Cette diminution simultanée de l’activité d’AP-1 et du niveau de miR-155 pourrait se révéler intéressante pour une possible application thérapeutique du resvératrol comme agent anti-inflammatoire et anticancéreux.
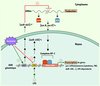 |
Figure 1 Le resvératrol module l’expression de micro-ARN. Dans la lignée de monocytes humains THP-1, le resvératrol diminue le niveau de miR-155 via la surexpression de miR-663. miR-663 cible les protéines JunB et JunD [5] et module ainsi l’expression du complexe de transcription pro-inflammatoire AP-1. LPS : lipopolysaccharide bactérien ; 155 : gène codant le miARN 155 non traductible ; 663 : gène codant le miARN 663 non traductible ; Jun B, Jun D : protéines constituant le complexe AP-1, stimulateur de la transcription de gènes pro-inflammatoires ; gène cible : gène pro-inflammatoire. |
La deuxième étude [8] a démontré, dans une lignée de cellules tumorales colorectales humaines (SW480), que le resvératrol entraîne aussi, de manière dépendante ou non de la surexpression de miR-663, la diminution du transcrit du gène codant le TGFβ1 (transforming growth factor β1). Par ailleurs, le criblage du profil de miARN à l’aide de puces montre que d’autres miARN oncogéniques sont réprimés : par exemple ceux qui ciblent les transcrits des gènes DICER1 (la RNase III impliquée dans la maturation des miARN), PDCD4 (programmed cell death protein 4) et PTEN (phosphatase and tensin homolog), des effecteurs-clés de la voie de signalisation du TGFβ. Le TGFβ, lorsqu’il se fixe sur ses récepteurs, induit une des voies de signalisation cellulaires fréquemment altérées dans les cancers [9]. L’expression du TGFβ est élevée dans les cancers colorectaux métastatiques et est corrélée avec le grade de la tumeur. Comme le montre la Figure 2, le TGFβ1 a un double effet en fonction du grade de la tumeur : suppresseur de tumeur au stade prétumoral et promoteur de métastases à un stade avancé malin. Puisque le resvératrol entraîne une modification de la population cellulaire de miARN de type oncogènes ou suppresseurs de tumeurs, une voie de recherche intéressante pourrait exploiter ces données sur un plan thérapeutique, en fonction du grade de la tumeur.
 |
Figure 2 Effet du resvératrol dans les cellules de la lignée tumorale humaine SW48. Le resvératrol module le niveau de microARN en ciblant des gènes codant des suppresseurs de tumeur et des effecteurs de la signalisation de TGFβ1 [9]. |
Conflit d’intérêts
Les auteurs déclarent n’avoir aucun conflit d’intérêts concernant les données publiées dans cet article.
Références
- Lee RC, Feinbaum RL, Ambros V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993 ; 75 : 843-866. [CrossRef] [PubMed] [Google Scholar]
- Calin GA, Sevignani C, Dumitru CD, et al. Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers. Proc Natl Acad Sci USA 2004 ; 101 : 2999-3004. [CrossRef] [Google Scholar]
- Parker JA, Arango M, Abderrahmane S, et al. Neuroprotection par l’activation des sirtuines dans des modèles simplifiés de chorée de Huntington. Med Sci (Paris) 2005 ; 21 : 556-557. [PubMed] [Google Scholar]
- Delmas D, Lançon A, Colin D, et al. Resveratrol as chemopreventive agent : new concepts to put forward the molecule to fight cancer. Curr Drug Targets 2006 ; 7 : 423-442. [Google Scholar]
- Tili E, Michaille JJ, Adair B, et al. Resveratrol decreases the levels of miR-155 by upregulating miR-663, a microRNA targeting JunB and JunD. Carcinogenesis 2010 ; 31 : 1561-1566. [CrossRef] [PubMed] [Google Scholar]
- Udenigwe CC, Ramprasath VR, Aluko RE, Jones PJ. Potential of resveratrol in anticancer and anti-inflammatory therapy. Nutr Rev 2008 ; 66 : 445-454. [CrossRef] [PubMed] [Google Scholar]
- Ruggiero T, Trabucchi M, De Santa F, et al. LPS induces KH-type splicing regulatory protein-dependent processing of microRNA-155 precursors in macrophages. Faseb J 2009 ; 23 : 2898-2908. [CrossRef] [PubMed] [Google Scholar]
- Tili E, Michaille JJ, Alder H, et al. Resveratrol modulates the levels of microRNAs targeting genes encoding tumor-suppressors and effectors of TGF beta signaling pathway in SW480 cells. Biochem Pharmacol 2010 ; 80 : 2057-2065. [CrossRef] [PubMed] [Google Scholar]
- Akhurst RJ. TGF beta signaling in health and disease. Nat Genet 2004 ; 36 : 790-792. [CrossRef] [PubMed] [Google Scholar]
© 2011 médecine/sciences – Inserm / SRMS
Liste des figures
|
Figure 1 Le resvératrol module l’expression de micro-ARN. Dans la lignée de monocytes humains THP-1, le resvératrol diminue le niveau de miR-155 via la surexpression de miR-663. miR-663 cible les protéines JunB et JunD [5] et module ainsi l’expression du complexe de transcription pro-inflammatoire AP-1. LPS : lipopolysaccharide bactérien ; 155 : gène codant le miARN 155 non traductible ; 663 : gène codant le miARN 663 non traductible ; Jun B, Jun D : protéines constituant le complexe AP-1, stimulateur de la transcription de gènes pro-inflammatoires ; gène cible : gène pro-inflammatoire. |
| Dans le texte | |
|
Figure 2 Effet du resvératrol dans les cellules de la lignée tumorale humaine SW48. Le resvératrol module le niveau de microARN en ciblant des gènes codant des suppresseurs de tumeur et des effecteurs de la signalisation de TGFβ1 [9]. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.