")
")
| Issue |
Med Sci (Paris)
Volume 22, Number 3, Mars 2006
|
|
|---|---|---|
| Page(s) | 239 - 240 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/2006223239 | |
| Published online | 15 March 2006 | |
Mutations hypomorphiques de RAG1 et infections à CMV : un nouveau phénotype de déficit immunitaire
Hypomorphic RAG1 mutations and CMV infection: a new phenotype of severe combined immunodeficiency
1
Développement normal et pathologique du système immunitaire, Inserm U429, Hôpital Necker-Enfants Malades, 149, rue de Sèvres, 75015 Paris, France
2
Département de microbiologie et d’immunologie, Hôpital Sainte Justine, 3175, côte Sainte Catherine, Montréal, Québec, H3T 1C5 Canada
3
Développement normal et pathologique du système immunitaire, Inserm U429, Université René Descartes, Faculté de médecine, AP-HP, Hôpital Necker-Enfants Malades, 149, rue de Sèvres, 75015 Paris, France
4
Unité d’immunité antivirale, de biothérapie et de vaccination, Institut Pasteur, 27, rue du Docteur Roux, 75015 Paris, France
5
UMR CNRS 5164, CIRID, Université de Bordeaux, 146, rue Léo Saignat, BP14, 33076 Bordeaux Cedex, France
*
This email address is being protected from spambots. You need JavaScript enabled to view it.
Les déficits immunitaires combinés sévères (DICS) consistent en des défauts complets génétiquement déterminés de l’immunité adaptative. À ce jour, douze mécanismes ont été identifiés et rendent compte de plus de 90 % des cas. Un défaut de réarrangement somatique des gènes des récepteurs de l’antigène des lymphocytes T (TCR) et des lymphocytes B (BCR), étape indispensable à la différenciation et à la diversification des lymphocytes T et B, conduit à l’absence des lymphocytes T associée à l’absence de lymphocytes B contrastant avec la présence normale de cellules NK [DICS T(-) B(-) NK (+)]. Des mutations dans les gènes codant RAG1, RAG2 ou Artémis, protéines indispensables au processus de réarrangement, sont à l’origine de ce type de DICS [1, 2]. Cependant, des mutations hypomorphes de ces gènes peuvent, dans certains cas, permettre une différenciation résiduelle de lymphocytes T et/ou B conduisant à des phénotypes variables [1, 3]. Ces DICS « atypiques » diffèrent par la présence de lymphocytes T. Ainsi, le syndrome d’Omenn est caractérisé par l’expansion de lymphocytes T oligoclonaux, autoréactifs, qui infiltrent, en particulier, la peau et la muqueuse intestinale [1, 4].
Nous avons récemment identifié des mutations hypomorphes de RAG1 chez quatre patients qui présentaient un tableau clinique particulier associant une infection grave à cytomégalovirus (CMV) et des manifestation auto-immunes de type anémie hémolytique et neutropénie [5]. Un 5e cas a également été séparément décrit [6]. Chez ces patients, des lymphocytes T et NK étaient détectables dans le sang périphérique. Chez trois d’entre eux, des lymphocytes B étaient également détectables dans le sang, associés à la présence d’immunoglobulines sériques et d’anticorps notamment dirigés contre le CMV. Les manifestations auto-immunes, anémie et neutropénie étaient liées à la présence d’auto-anticorps. Globalement, le phénotype de ces patients apparaît comme assez éloigné de celui des DICS T(-) B(-) NK(+) classiquement caractérisé par l’absence complète de lymphocytes T, de lymphocytes B et d’immunoglobulines sériques. Cependant, l’analyse phénotypique des lymphocytes T circulants révéla que la majorité TCR de type γ/δ, récepteur normalement exprimé par moins de 15 % des lymphocytes T du sang, et que seuls, 8 à 30 % des lymphocytes T des patients exprimaient un TCR α/β. De plus, ces derniers lymphocytes T présentaient un phénotype mémoire, [CD45RO], et exprimaient des TCR α/β dont la diversité était restreinte. L’ensemble de ces observations a suggéré l’existence suggéré l’existence d’un défaut de recombinaison des gènes des TCR. L’infection d’un défaut de recombinaison des gènes des TCR. L’infection par le CMV, pourrait avoir induit l’expansion des lymphocytes T TCR γ/δ dans un contexte de restriction d’hébergement du répertoire des lymphocytes T TCR γ/δ. En effet, une expansion de certains lymphocytes T TCR γ/δ est observée chez les patients infectés par le CMV au cours d’un traitement immunosuppresseur nécessité par une greffe de rein. Ces lymphocytes T TCR γ/δ exercent une activité cytotoxique à l’égard de cellules infectées par le CMV et sécrètent spécifiquement du TNF [7]. Chez les patients que nous décrivons, une telle activité n’a pas pu être mise en évidence in vitro, mais elle peut exister comme l’a montré Ehl et al. dans un cas similaire [6].
L’auto-immunité dépendante des anticorps observée chez trois patients pourrait être le simple témoin d’un défaut d’homéostasie lymphocytaire dans un contexte de déficit immunitaire. Des hypothèses alternatives impliquent un rôle dans le déclenchement de l’auto-immunité à l’infection par le CMV souvent associée à des cytopénies auto-immunes [8] et/ou aux lymphocytes T TCR γ/δ dont le rôle dans l’auto-immunité est évoqué [9].
Ces quatre patients présentent une mutation homozygote de RAG1, compatible avec une activité protéique résiduelle. Chez deux patients, il s’agit d’une mutation conduisant à une substitution d’acide aminé. Dans les deux autres cas, il s’agit de délétions conduisant à des codons stop ; cependant, la présence de différents sites d’initiation de la traduction permet alors la synthèse d’une protéine tronquée dans sa partie amino-terminale et partiellement fonctionnelle. Ces mutations sont soit similaires soit identiques à des mutations observées chez des patients atteints du syndrome d’Omenn, et dont l’activité résiduelle de la protéine a été démontrée [1, 10].
La description d’un nouveau tableau clinico-biologique associé à des mutations hypomorphes de RAG1 élargit le spectre d’expression de ce déficit. Ainsi, aussi bien des facteurs environnementaux, tels qu’une infection à CMV, et/ou génétiques - encore indéterminés dans le cas du syndrome d’Omenn - en modifient l’expression (Figure 1).
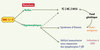 |
Figure 1. Spectre d’expression phénotypique associé aux mutations des gènes codant les protéines Rag-1 et Rag-2. |
Il est très vraisemblable que cette diversité d’expression est également rencontrée dans d’autres anomalies héréditaires monogéniques du système immunitaire, un concept largement extensible à l’ensemble des maladies héréditaires monogéniques.
Références
- Corneo B, Moshous D, Gungor T, et al. Identical mutations in RAG1 or RAG2 genes leading to defective V(D)J recombinase activity can cause either T-B-severe combined immune deficiency or Omenn syndrome. Blood 2001; 97 : 2772–6. [Google Scholar]
- Moshous D, Callebaut I, de Chasseval R, et al. Artemis, a novel DNA double-strand break repair/V(D)J recombination protein, is mutated in human severe combined immune deficiency. Cell 2001; 105 : 177–86. [Google Scholar]
- Ege M, Ma Y, Manfras B, et al. Omenn syndrome due to Artemis mutations. Blood 2005; 105 : 4179–86. [Google Scholar]
- Rieux-Laucat F, Bahadoran P, Brousse N, et al. Highly restricted human T cell repertoire in peripheral blood and tissue-infiltrating lymphocytes in Omenn’s syndrome. J Clin Invest 1998; 102 : 312–21. [Google Scholar]
- De Villartay JP, Lim A, Al-Mousa H, et al. A novel immunodeficiency associated with hypomorphic RAG1 mutations and CMV infection. J Clin Invest2005; 115 : 3291–9. [Google Scholar]
- Ehl S, Schwarz K, Enders A, et al. A variant of SCID with specific immune responses and predominance of gammadelta T cells. J Clin Invest 2005; 115 : 3140–8. [Google Scholar]
- Dechanet J, Merville P, Lim A, et al. Implication of gammadelta T cells in the human immune response to cytomegalovirus. J Clin Invest 1999; 103 : 1437–49. [Google Scholar]
- Murray JC, Bernini JC, Bijou HL, et al. Infantile cytomegalovirus-associated autoimmune hemolytic anemia. J Pediatr Hematol Oncol 2001; 23 : 318–20. [Google Scholar]
- Ware RE, Howard TA. Elevated numbers of gamma-delta (gamma delta+) T lymphocytes in children with immune thrombocytopenic purpura. J Clin Immunol 1994; 14 : 237–47. [Google Scholar]
- Villa A, Sobacchi C, Vezzoni P. Recombination activating gene and its defects. Curr Opin Allergy Clin Immunol 2001; 1 : 491–5. [Google Scholar]
© 2006 médecine/sciences - Inserm / SRMS
Liste des figures
|
Figure 1. Spectre d’expression phénotypique associé aux mutations des gènes codant les protéines Rag-1 et Rag-2. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.