")
")
| Issue |
Med Sci (Paris)
Volume 21, Number 3, Mars 2005
|
|
|---|---|---|
| Page(s) | 309 - 314 | |
| Section | M/S revues | |
| DOI | https://doi.org/10.1051/medsci/2005213309 | |
| Published online | 15 March 2005 | |
Peptides transducteurs
La face utile d’un nouveau mode de signalisation
Transduction peptides, the useful face of a new signaling mechanism
1
Équipe biologie cellulaire des homéoprotéines
2
Équipe développement et neuropharmacologie. CNRS UMR 8542, École Normale Supérieure, 46, rue d’Ulm, 75230 Paris Cedex 05, France
*
This email address is being protected from spambots. You need JavaScript enabled to view it.
*
This email address is being protected from spambots. You need JavaScript enabled to view it.
Résumé
En 1988, A.D. Frankel et C.O. Pabo proposent que le facteur de transcription TAT du virus de l’immunodéficience humaine (VIH) est internalisé par les cellules. Parallèlement, des travaux développés dans notre laboratoire démontrent que le domaine de fixation sur l’ADN (l’homéodomaine) d’un facteur de transcription de la famille des homéoprotéines passe directement de l’extérieur vers l’intérieur de la cellule. La séquence responsable de cette internalisation est la troisième hélice de l’homéodomaine, longue de 16 acides aminés. La conservation de cette structure dans tous les facteurs de transcription de la famille des homéoprotéines suggère que l’internalisation est une propriété partagée par la grande majorité de ces protéines. Ces études ont ouvert la voie au concept physiologique de protéine messagère et au développement de peptides de transduction. Ces peptides sont utilisés comme vecteurs pour internaliser dans les cellules, in vitro et in vivo, des cargos hydrophiles de composition et de structure très variées. Cet article fait le point sur les mécanismes impliqués, les utilisations vérifiées ou potentielles de ces peptides et les difficultés rencontrées dans certaines de leurs utilisations, sans oublier les implications physiologiques de ce phénomène.
Abstract
Tranduction peptides that cross the plasma membrane of live cells are commonly used for the in vitro and in vivo targeting of hydrophilic drugs into the cell interior. Although this family of peptides has recently increased and will probably continue to do so, the two mainly used peptides are derived from transcription factors. Indeed, TAT is a 12 amino acid long arginine-rich peptide present in the HIV transcription factor, and penetratin - or its variants - corresponds to 16 amino acids that define the highly conserved third helix of the DNA-binding domain (homeodomain) of homeoprotein transcription factors. In this review, we shall recall the different steps that have led to the discovery of transduction peptides and present the most likely hypotheses concerning the mechanisms involved in their internalization. At the risk of being incomplete or, even, biased, we shall concentrate on penetratins and TAT. The reason is that these peptides have been studied for over ten years leading to the edification of robust knowledge regarding their properties. This attitude will not preclude comparisons with other peptides, if necessary. Our goal is to describe the mode of action of these transduction peptides, their range of activity in term of cell types that accept them and cargoes that they can transport, and, also, some of the limitations that one can encounter in their use. Finally, based on the idea that peptide transduction is the technological face of a physiological property of some transcription factors, we shall discuss the putative physiological function of homeoprotein transduction, and, as a consequence, the possibility to use these factors as therapeutic proteins.
© 2005 médecine/sciences - Inserm / SRMS
Transduction et protéines messagères
Par transduction, nous désignons la capacité d’une molécule de passer à travers une membrane plasmique et d’accéder aux compartiments cytosolique et nucléaire (non vésiculaires). Cette propriété est physiologiquement partagée par de nombreux agents lipophiles, par exemple les rétinoïdes, dont l’entrée dans la cellule et la fixation à un récepteur nucléaire a des conséquences fonctionnelles au niveau de la transcription de gènes cibles. Le passage à travers les membranes du monoxyde d’azote (NO) est un autre exemple de transduction.
La transduction protéique pose le problème de la perméabilité de la membrane plasmique vis-à-vis d’une molécule hydrophile. Certes, nous connaissons des peptides qui traversent les membranes, comme les défensines, peptides antibiotiques d’insectes, ou certaines toxines, comme la toxine diphtérique. Mais, dans ces deux exemples, le passage implique la formation d’un canal formé par l’insertion de peptides amphiphiles dans le plan perpendiculaire à celui de la bicouche lipidique, ménageant un couloir hydrophile aux autres peptides (Figure 1A). Ce pore peut être formé au niveau de la membrane plasmique (cas des défensines), ou nécessiter un changement de conformation catalysé par le pH acide du milieu intravésiculaire. Dans un cas comme dans l’autre, nous avons affaire à un processus, certes physiologique, mais dont le résultat peut entraîner la mort de la cellule concernée.
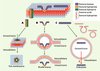 |
Figure 1. Modes d’internalisation des peptides. Les mécanismes d’internalisation des peptides peuvent faire appel (A) à la formation d’un canal dans la membrane plasmique, (B) à la formation de micelles inverses (cas des peptides dotés d’une structure de type penetratin) ou (C) à l’endocytose, celle-ci étant probablement suivie d’une rupture de la vésicule. |
C’est d’ailleurs aussi dans un contexte pathologique que fut observée, en 1988, l’internalisation de la protéine TAT du VIH [1]. Le transfert de facteurs de transcription fut évoqué de nouveau quelques années plus tard, à partir de 1991, à propos des protéines à homéodomaine - ou homéoprotéines -, une classe de facteurs de transcription à activité morphogénétique.
La penetratin et ses mécanismes d’internalisation
En 1991, c’est pour tester le rôle possible des homéoprotéines dans la morphogenèse des neurones murins que l’homéodomaine d’Antennapedia (une homéoprotéine de drosophile) fut introduit, mécaniquement, dans des neurones en culture. Les 60 acides aminés de l’homédomaine étant largement conservés, l’idée était d’utiliser ce domaine de fixation à l’ADN comme antagoniste compétitif des homéoprotéines endogènes. Cette expérience, prouvant le rôle de cette classe de facteurs de transcription dans la morphogenèse des cellules nerveuses postmitotiques, mit aussi en évidence, au cours d’un contrôle, l’internalisation spontanée de l’homéodomaine dans les cellules [2].
Il fut alors démontré que l’entrée de l’homéodomaine pouvait être observée à basse température, c’est-à-dire dans des conditions excluant toute endocytose, dans tous les types cellulaires et toutes les cellules de la culture. Par ailleurs, la troisième hélice de l’homéodomaine, longue de seulement 16 acides aminés, conservait l’ensemble de ces propriétés [3]. Cette séquence, baptisée penetratin, se structure en hélice α amphiphile en milieu hydrophobe, avec une face basique, riche en lysines et en arginines, et une face hydrophobe [3].
La courte taille du peptide permit, par une étude « structure-fonction », d’identifier les acides aminés indispensables à l’internalisation et de produire une série importante de peptides transducteurs dérivés de la penetratin (Tableau I) [4]. Trois résultats sont particulièrement importants : le premier est le rôle clé joué par le tryptophane en position 48 (W48) de l’homéodomaine, qui ne peut être remplacé par aucun autre acide aminé hydrophobe sans diminution drastique de la transduction ; le second est l’internalisation du peptide inverse ou d’une penetratin composée d’acides aminés dextrogyres, validant l’absence de récepteur chiral ; le troisième, enfin, démontré par l’introduction de résidus proline (Tableau I), est la « dispensabilité » d’une structure en hélice α.
Peptides transducteurs. * Dérivé de la penetratin non internalisé.
La comparaison de l’interaction, avec les phospholipides membranaires, de peptides actifs et de variants inactifs, notamment du variant W48F (dans lequel le tryptophane 48 a été remplacé par une phénylalanine), a conduit à proposer un mécanisme d’entrée par formation de micelles inverses. Bien que ces micelles n’aient pas été observées directement, ce modèle est corroboré par les expériences menées par des chimistes et des physiciens ayant étudié le mécanisme d’internalisation [5] ; l’interaction des peptides avec des bicouches lipidiques a démontré l’importance des charges négatives portées par certains phospholipides [6]. Ces données ont conduit au modèle de transduction illustré dans la Figure 1B. Cette même figure schématise les autres modes d’internalisation possibles utilisés par certains peptides, comme la formation d’un canal dans la membrane plasmique (Figure 1A) ou l’endocytose (Figure 1C).
Autres peptides de transduction
Peu après la découverte des propriétés de transduction de la penetratin et de ses dérivés, un peptide de 11 acides aminés, probablement responsable de la transduction de TAT, fut identifié dans sa région carboxyterminale [7, 8]. S’ensuivit la synthèse de peptides absents des protéines naturellement exprimées par les organismes, mais dont les propriétés physico-chimiques prédisaient des propriétés de transduction (Tableau I). Les peptides strictement amphipatiques (MAP), ou cationiques (11R), sont des exemples extrêmes. La plupart de ces peptides ont été utilisés comme vecteurs peptidiques pour l’adressage d’agents hydrophiles dans les cellules [9] et ont fait l’objet, comme le peptide TAT et la penetratin, d’études mécanistiques.
Les mécanismes d’entrée de ces différents peptides dans la cellule restent mal définis, mais on peut exclure pour certains d’entre eux un mode d’entrée semblable à celui des penetratins. Pour ce qui est de TAT, l’équipe de S.F. Dowdy a récemment apporté les preuves d’une entrée par endocytose, suivie d’une rupture de la vésicule favorisant le passage dans le cytoplasme ; de fait, l’ajout d’un peptide fusogène en position carboxyterminale de TAT augmente ses capacités transductrices [10]. On peut donc avancer que TAT (ou les autres peptides poly-arginines) et les penetratins entrent dans la cellule par deux mécanismes différents. Pour les autres peptides transducteurs, il serait imprudent de formuler des hypothèses trop précises à ce stade.
Il faut également garder à l’esprit qu’un même peptide de transduction pourrait entrer dans les cellules par des mécanismes distincts. Par exemple, l’internalisation de la penetratin à 4 °C, sans endocytose, n’exclut pas qu’elle puisse aussi entrer par endocytose. Plus important encore, le vecteur pourrait, une fois lié à un cargo, adopter des propriétés nouvelles résultant des caractéristiques combinées du vecteur et du cargo, et susceptibles d’affecter l’efficacité et le mode d’internalisation.
Vectorisation par les peptides de transduction
Au-delà des possibles applications dans le domaine thérapeutique, les peptides transducteurs ont été utilisés de façon répétée pour l’étude de nombreuses fonctions physiologiques. À titre d’exemple, la Figure 2 illustre quelques cas d’internalisation in vitro et in vivo de la penetratin1. La nature des cargos ainsi internalisés est très variée : oligonucléotides antisens, antisens peptidiques (PNA), petits ARN interférents (ARNsi), peptides ou phosphopeptides, protéines entières.
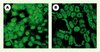 |
Figure 2. Internalisation de la penetratin ex vivo et in vivo.A. La penetratin couplée à un fluorochrome est internalisée par l’ensemble des cellules (lignée MDCK) et s’accumule dans le noyau et le cytoplasme. B.. Détection du même peptide dans le rein, après injection intrapéritonéale chez la souris. |
L’avantage de cette technique, par rapport aux stratégies fondées sur l’expression de la molécule d’intérêt par la cellule, est le peu de toxicité des vecteurs, leur efficacité in vitro et in vivo et la possibilité de contrôler la quantité de cargo internalisé : cette dernière caractéristique, notamment, évite les surexpressions massives qui accompagnent une transfection ou une infection virale, et permet de travailler dans des conditions quasi physiologiques.
Certains points positifs ont cependant leur revers, comme l’entrée dans tous les types cellulaires et dans la totalité des cellules, ce qui rend difficile un adressage cellulaire spécifique s’il n’y a pas eu ajout de domaines d’adressage. Un autre point qui a ses bons et ses mauvais côtés est le caractère transitoire de l’effet biologique, puisque les cargos ne modifient pas le génome de la cellule et ont, par eux-mêmes, une demi-vie limitée. Dans certains cas, cependant, cette expression transitoire peut avoir des effets définitifs, comme l’induction de l’apoptose ou l’engagement dans une voie de différenciation.
L’entrée des cargos peut se produire sans liaison covalente avec le vecteur. Ce procédé, popularisé sous le terme de « chariot », est d’utilisation aisée, mais demande un rapport vecteur/cargo élevé, jusqu’à 100 molécules de vecteur par molécule de cargo [11]. Même si ces vecteurs sont peu toxiques, un tel envahissement de la cellule par des vecteurs le plus souvent cationiques peut ne pas être sans conséquences sur la physiologie cellulaire. C’est pourquoi la vectorisation par formation d’une liaison covalente entre le vecteur et le cargo semble préférable. Cette liaison peut être irréversible, comme dans le cas d’une protéine de fusion portant un domaine de transduction. Elle peut aussi être réversible, quand le vecteur et le cargo sont liés par un pont disulfure, dont la demi-vie est brève dans le milieu réducteur intracellulaire. Dans ce cas, le cargo est libéré et son affinité pour une cible détermine sa localisation finale. En revanche, si le lien est non labile, la localisation dépend des propriétés de la chimère cargo-vecteur. Même si, dans de nombreux cas, la localisation reflète l’interaction du cargo avec sa cible, on ne peut cependant exclure une influence du vecteur sur la distribution subcellulaire. Évidemment, des séquences d’adressage peuvent être ajoutées, qui rapprocheraient le composé du compartiment souhaité : une séquence de localisation nucléaire (NLS) pour adressage nucléaire, une séquence CAAX pour accrochage à la membrane…
Une question récurrente est celle de la taille du cargo. Pour ce qui est de la penetratin ou de TAT, des cargos protéiques composés de quelques acides aminés à plusieurs centaines de résidus ont été internalisés avec succès. Des échecs sont possibles, même pour des polypeptides relativement courts, mais il semble que ce soit là un problème de structure plutôt que de taille. Pour un polypeptide, en-dessous de 100 acides aminés, aucun échec n’a été rapporté ; pour des tailles supérieures, en l’absence actuelle de règle prédictive générale, la seule solution est d’essayer. Quant à la dénaturation des protéines nécessaire à leur internalisation, proposée dans les premiers travaux sur TAT, c’est là une option rendue obsolète par les travaux récents de l’équipe de S.F. Dowdy qui démontrent que l’entrée s’effectue par endocytose, suivie d’une rupture de la vésicule [10].
Signification physiologique du phénomène de transduction protéique
Il est significatif que les deux vecteurs peptidiques les plus utilisés à ce jour sont dérivés de protéines de la famille des facteurs de transcription douées d’une propriété de transfert intercellulaire. Pour la protéine TAT, après l’observation initiale de A.D. Frankel et de ses collaborateurs [1], sa capacité de transactivation transcellulaire a été démontrée en culture cellulaire [12, 13], les conséquences physiopathologiques de ce transfert demandant à être élucidées.
Pour ce qui est des protéines à homéodomaine, leur sécrétion comme leur internalisation ont été démontrées in vitro, et le mécanisme de leur sécrétion partiellement identifié [14]. Les homéoprotéines ne possèdent pas de séquence d’adressage à un compartiment classique de sécrétion impliquant l’appareil de Golgi. De façon surprenante, la sécrétion se fait par un mécanisme distinct du mécanisme d’internalisation : cela est démontré par l’existence de mutants d’internalisation toujours capables d’être sécrétés, et par le caractère vésiculaire, et dépendant de l’énergie, de la sécrétion. Une séquence de sécrétion a été identifiée dans l’homéodomaine, dont les caractéristiques sont celles d’une séquence d’export nucléaire [15]. Par ailleurs, la sécrétion est réglée par la phosphorylation d’un domaine riche en sérines localisé, pour l’homéoprotéine Engrailed (impliquée dans la formation du cervelet et du mésencéphale postérieur, ainsi que dans le guidage axonal), en amont de son homéodomaine [16] (Figure 3).
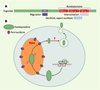 |
Figure 3. Processus de sécrétion proposé pour l’homéoprotéine Engrailed.A. Localisation des séquences permettant l’internalisation et la sécrétion. B. Processus de sécrétion. La protéine synthétisée dans la cellule donneuse doit passer par le noyau pour, après export nucléaire, être adressée à des vésicules de sécrétion ayant les caractéristiques de cavéoles (richesse en glycosphingolipides et cholestérol) : la capacité d’adressage vésiculaire et de sécrétion de la protéine est en effet strictement corrélée à l’accumulation de cette dernière dans le noyau. Il est particulièrement important de constater qu’in vivo, les protéines Engrailed-1 et Engrailed-2 ont été retrouvées dans ce même compartiment de sécrétion (en continuité avec le milieu extracellulaire, après fusion de la vésicule avec la membrane plasmique). |
Une fonction physiologique importante de ce mode de transduction du signal est suggérée par des travaux récemment réalisés sur le transport axonal antérograde d’homéoprotéines [17], par leur activité trans-synaptique sur la maturation nerveuse [18] et par leur interaction avec le facteur d’initiation de la traduction eIF4E, in vitro comme in vivo [17, 19]. Ces travaux rejoignent ainsi des observations plus anciennes sur l’homéoprotéine Bicoïd, dont l’interaction avec eIF4E inhibe la traduction de Caudal [20]. Il est important de noter que le domaine d’interaction avec eIF4E, fonctionnel dans Bicoïd, a été retrouvé dans 200 homéoprotéines [19].
Conclusions
De ces observations récentes sur les peptides de transduction, nous tirons deux conclusions. La première est que les homéoprotéines, dont nombre sont impliquées dans des maladies, doivent être considérées comme des protéines thérapeutiques potentielles. La faisabilité de cette approche a récemment été démontrée par l’utilisation de la protéine à homéodomaine HOXB4 comme protéine thérapeutique dans l’amplification de cellules souches hématopoïétiques [21] (→). Une deuxième conclusion est que, derrière les propriétés de la penetratin et de ses dérivés actifs, se cache probablement un mode insoupçonné de signalisation cellulaire par régulation non autonome (la protéine transférée n’étant pas synthétisée par la cellule receveuse) de l’expression génique, aux niveaux transcriptionnel et traductionnel. L’intérêt de cette régulation non autonome (Figure 4) est lié à la fonction même de ces protéines comme marqueurs de position : la réponse de la cellule receveuse serait alors modulée en fonction de l’origine topologique de la cellule donneuse. Pour prendre le cas d’un réseau de neurones, la cellule recevant un neuromédiateur recevrait également une information sur la position, dans le réseau, du corps cellulaire dont la terminaison libère le neuromédiateur. Une prédiction raisonnable - étayée par l’expression persistante de ces facteurs chez l’adulte - est que la compréhension de ce mode de signalisation aura des implications en pathologie.
(→) m/s 2004, n° 11, p. 959
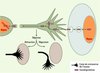 |
Figure 4. Rôle potentiel des homéoprotéines dans la navigation des cônes de croissances axonaux. Les homéoprotéines sécrétées par les cellules situées à proximité d’un cône de croissance sont internalisées par ce dernier. Elles peuvent régler la traduction localement au niveau du cône de croissance ou être transportées vers le noyau de l’axone. La nature de l’homéoprotéine internalisée et l’origine anatomique du cône déterminent la réponse du cône de croissance (attraction ou répulsion). |
Il est impossible de donner ici un compte rendu exhaustif des utilisations publiées ; le lecteur intéressé pourra facilement les trouver sur la toile à l’aide de certains mots-clés comme penetratin, antennapedia, TAT, cell permeable peptides, transduction peptides.
Références
- Frankel AD, Pabo CO. Cellular uptake of the Tat protein from human immunodeficiency virus. Cell 1988; 55 : 1189–93. [Google Scholar]
- Joliot A, Pernelle C, Deagostini-Bazin H, et al. Antennapedia homeobox peptide regulates neural morphogenesis. Proc Natl Acad Sci USA 1991; 88 : 1864–8. [Google Scholar]
- Derossi D, Joliot AH, Chassaing G, et al. The third helix of Antennapedia homeodomain translocates through biological membranes. J Biol Chem 1994; 269 : 10444–50. [Google Scholar]
- Derossi D, Calvet S, Trembleau A, et al. Cell internalization of the third helix of the Antennapedia homeodomain is receptor-independent. J Biol Chem 1996; 271 : 18188–93. [Google Scholar]
- Berlose JP, Convert O, Derossi D, et al. Conformational and associative behaviours of the third helix of Antennapedia homeodomain in membrane-mimetic environments. Eur J Biochem 1996; 242 : 372–86. [Google Scholar]
- Christiaens B, Symoens S, Verheyden S, et al. Tryptophan fluorescence study of the interaction of penetratin peptides with model membranes. Eur J Biochem 2002; 269 : 2918–26. [Google Scholar]
- Fawell S, Seery J, Daikh Y, et al. Tat-mediated delivery of heterologous proteins into cells. Proc Natl Acad Sci USA 1994; 91 : 664–8. [Google Scholar]
- Anderson DC, Nichols E, Manger R, et al. Tumor cell retention of antibody Fab fragments is enhanced by an attached HIV TAT protein-derived peptide. Biochem Biophys Res Commun 1993; 194 : 876–84. [Google Scholar]
- Dupont E, Joliot A, Prochiantz A. Cell penetrating peptides. In : Langel U, ed. Penetratins. Boca Raton, Florida : CRC Press, 2002 : 23–51. [Google Scholar]
- Wadia JS, Stan RV, Dowdy SF. Transducible TAT-HA fusogenic peptide enhances escape of TAT-fusion proteins after lipid raft macropinocytosis. Nat Med 2004; 10 : 310–5. [Google Scholar]
- Dom G, Shaw-Jackson C, Matis C, et al. Cellular uptake of Antennapedia penetratin peptides is a two-step process in which phase transfer precedes a tryptophan-dependent translocation. Nucleic Acids Res 2003; 31 : 556–61. [Google Scholar]
- Helland DE, Welles JL, Caputo A, et al. Transcellular transactivation by the human immunodeficiency virus type 1 tat protein. J Virol 1991; 65 : 4547–9. [Google Scholar]
- Thomas CA, Dobkin J, Weinberger OK. TAT-mediated transcellular activation of HIV-1 long terminal repeat directed gene expression by HIV-1-infected peripheral blood mononuclear cells. J Immunol 1994; 153 : 3831–9. [Google Scholar]
- Joliot A, Maizel A, Rosenberg D, et al. Identification of a signal sequence necessary for the unconventional secretion of Engrailed homeoprotein. Curr Biol 1998; 8 : 856–63. [Google Scholar]
- Maizel A, Bensaude O, Prochiantz A, et al. A short region of its homeodomain is necessary for Engrailed nuclear export and secretion. Development 1999; 126 : 3183–90. [Google Scholar]
- Maizel A, Tassetto M, Filhol O, et al. Engrailed homeoprotein secretion is a regulated process. Development 2002; 129 : 3545–53. [Google Scholar]
- Nedelec S, Foucher I, Brunet I, et al. Emx2 homeodomain transcription factor interacts with eukaryotic translation initiation factor 4E (eIF4E) in the axons of olfactory sensory neurons. Proc Natl Acad Sci USA 2004; 101 : 10815–20. [Google Scholar]
- Sugiyama S, Volovitch M, Prochiantz A, et al. Specification of critical period plasticity by Otx2 homeoprotein in the visual pathway. Washington DC : Society for Neurosciences, 2004. [Google Scholar]
- Topisirovic I, Culjkovic B, Cohen N, et al. The proline-rich homeodomain protein, PRH, is a tissue-specific inhibitor of eIF4E-dependent cyclin D1 mRNA transport and growth. EMBO J 2003; 22 : 689–703. [Google Scholar]
- Niessing D, Blanke S, Jackle H. Bicoid associates with the 5’-cap-bound complex of caudal mRNA and represses translation. Genes Dev 2002; 16 : 2576–82. [Google Scholar]
- Amsellem S, Pflumio F, Bardinet D, et al. Ex vivo expansion of human hematopoietic stem cells by direct delivery of the HOXB4 homeoprotein. Nat Med 2003; 9 : 1423–7. [Google Scholar]
Liste des tableaux
Liste des figures
|
Figure 1. Modes d’internalisation des peptides. Les mécanismes d’internalisation des peptides peuvent faire appel (A) à la formation d’un canal dans la membrane plasmique, (B) à la formation de micelles inverses (cas des peptides dotés d’une structure de type penetratin) ou (C) à l’endocytose, celle-ci étant probablement suivie d’une rupture de la vésicule. |
| Dans le texte | |
|
Figure 2. Internalisation de la penetratin ex vivo et in vivo.A. La penetratin couplée à un fluorochrome est internalisée par l’ensemble des cellules (lignée MDCK) et s’accumule dans le noyau et le cytoplasme. B.. Détection du même peptide dans le rein, après injection intrapéritonéale chez la souris. |
| Dans le texte | |
|
Figure 3. Processus de sécrétion proposé pour l’homéoprotéine Engrailed.A. Localisation des séquences permettant l’internalisation et la sécrétion. B. Processus de sécrétion. La protéine synthétisée dans la cellule donneuse doit passer par le noyau pour, après export nucléaire, être adressée à des vésicules de sécrétion ayant les caractéristiques de cavéoles (richesse en glycosphingolipides et cholestérol) : la capacité d’adressage vésiculaire et de sécrétion de la protéine est en effet strictement corrélée à l’accumulation de cette dernière dans le noyau. Il est particulièrement important de constater qu’in vivo, les protéines Engrailed-1 et Engrailed-2 ont été retrouvées dans ce même compartiment de sécrétion (en continuité avec le milieu extracellulaire, après fusion de la vésicule avec la membrane plasmique). |
| Dans le texte | |
|
Figure 4. Rôle potentiel des homéoprotéines dans la navigation des cônes de croissances axonaux. Les homéoprotéines sécrétées par les cellules situées à proximité d’un cône de croissance sont internalisées par ce dernier. Elles peuvent régler la traduction localement au niveau du cône de croissance ou être transportées vers le noyau de l’axone. La nature de l’homéoprotéine internalisée et l’origine anatomique du cône déterminent la réponse du cône de croissance (attraction ou répulsion). |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.