")
")
| Issue |
Med Sci (Paris)
Volume 42, Number 6-7, Juin-Juillet 2026
|
|
|---|---|---|
| Page(s) | 555 - 557 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/2026101 | |
| Published online | 17 juillet 2026 | |
Impact des nutriments sur l’efficacité de la thérapie anti-tumorale par blocage de points de contrôle immunitaire
Impact of nutrition of the efficacy of anti-tumoral therapy based on immune checkpoints blockade
Institut Curie, Inserm U932, Paris, France
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Les réponses immunitaires contre les tumeurs et les réponses aux traitements anti-cancéreux sont influencées par divers facteurs environnementaux et par le mode de vie [1]. La nutrition pourrait jouer un rôle important, mais les effets directs de différents nutriments sur les réponses immunitaires anti-tumorales restent mal compris.
La nutrition peut moduler le système immunitaire par le biais de métabolites produits soit directement lors de la digestion des aliments, soit par l’activité du microbiote intestinal. La diversité et la composition du microbiote intestinal jouent un rôle essentiel dans l’efficacité de l’immunothérapie par blocage des points de contrôle immunitaire [2]. Des études récentes sur des modèles murins ont également mis en évidence un rôle de certains nutriments dans les réponses immunitaires antitumorales. La vitamine C peut moduler le recrutement des lymphocytes T dans les tumeurs [3]. La vitamine E augmente l’efficacité du traitement par anticorps anti-PD1 ou anti-PDL1 en favorisant la fonction de présentation antigénique des cellules dendritiques [4]. De plus, certains métabolites produits par la cétogenèse hépatique augmentent les réponses immunitaires anti-tumorales en limitant l’expression des récepteurs inhibiteurs et en agissant en synergie avec le traitement anti-PD1 [5]. Ces résultats suggèrent que l’optimisation du régime alimentaire peut améliorer l’efficacité des traitements anti-cancéreux par blocage des points de contrôle immunitaire.
L’un des liens moléculaires entre l’alimentation et les cellules immunitaires est le récepteur d’aryl hydrocarbone (aryl hydrocarbon receptor, AhR), un facteur de transcription activé par ses ligands. AhR est synthétisé par de nombreuses cellules, en particulier les cellules épithéliales et certaines cellules immunitaires. Les ligands naturels de AhR sont principalement des indoles et des dérivés du tryptophane provenant de la nourriture ou du métabolisme du microbiote intestinal. Nous avons étudié l’impact des ligands alimentaires de AhR sur l’efficacité de l’immunothérapie anti-PD1 en utilisant un modèle murin [6].
Les ligands alimentaires de AhR sont essentiels à l’efficacité de l’immunothérapie anti-PD1
Nous avons utilisé des modèles tumoraux pré-cliniques chez des souris nourries avec un régime alimentaire synthétique, naturellement pauvre en ligands de AhR, ou avec le même régime supplémenté, à des doses physiologiques, en indole-3-carbinol, un ligand de AhR naturellement présent à haute dose dans les légumes de la famille des crucifères. Nous avons suivi la croissance tumorale chez des souris traitées ou non par anticorps anti-PD1 et nourries avec les différents régimes alimentaires. Nous avons constaté des réponses hétérogènes reflétant la situation chez les patients traités par ces anticorps (pas de réponse, réponse partielle avec ralentissement de la croissance tumorale, ou rejet de la tumeur). Néanmoins, l’effet du traitement était nettement moindre chez les souris nourries avec un régime pauvre en ligands de AhR. Nous avons obtenu les mêmes résultats pour trois modèles tumoraux différents : un fibrosarcome (MCA-101), une tumeur mammaire (E0771) et un mélanome (B16F10) [6]. Ces résultats suggèrent que les ligands alimentaires de AhR sont essentiels à l’efficacité de l’immunothérapie par anticorps anti-PD1.
La diminution des ligands de AhR dérivés du microbiote n’a aucun effet détectable sur l’efficacité de l’immunothérapie anti-PD1
Certaines bactéries du microbiote intestinal, en particulier les Lactobacilli, produisent également des ligands de AhR. Afin de déterminer si ces ligands étaient également importants pour l’efficacité du traitement anti-PD1, nous avons réalisé plusieurs séries d’expériences dans lesquelles nous avons diminué la production de ces ligands d’origine bactérienne. D’une part, nous avons éliminé les bactéries Lactobacilli en traitant les souris avec de l’ampicilline par voie orale. D’autre part, nous avons utilisé des souris axéniques (dépourvues de microbiote) dont nous avons ensemencé l’intestin, par voie orale, avec des bactéries E. Coli capables ou non de produire des indoles grâce à l’expression de l’enzyme tryptophanase. Dans ces expériences, nous n’avons constaté aucun effet de ces interventions sur l’efficacité du traitement anti-PD1 [6]. Nous avons ainsi montré que l’absence de différents ligands de AhR d’origine bactérienne n’avait pas d’effet notable sur l’efficacité de l’immunothérapie par anticorps anti-PD1.
L’absence des ligands alimentaires de AhR n’affecte pas le profil immunitaire tumoral de base
Afin d’étudier l’impact des ligands alimentaires de AhR sur l’infiltration des tumeurs par les cellules immunitaires, nous avons caractérisé les cellules immunitaires présentes dans la tumeur avant le traitement anti-PD1 par une analyse transcriptomique de cellules uniques (single-cell RNA-sequencing) couplée à la cytométrie en flux. L’infiltration des tumeurs par les monocytes, macrophages, cellules dendritiques, granulocytes neutrophiles, lymphocytes T CD4+ et CD8+ ou lymphocytes NK (natural killer) était identique chez des souris nourries avec le régime pauvre en ligands de AhR, ou avec le même régime supplémenté en indole-3-carbinol [6]. Nous avons donc émis l’hypothèse que les ligands alimentaires de AhR modulent l’efficacité du traitement en agissant sur la réponse à l’anticorps anti-PD1, plutôt que sur le profil immunitaire de base.
Pour vérifier cette hypothèse, nous avons ensuite analysé l’expression de plusieurs molécules effectrices du système immunitaire dans des tumeurs de souris nourries avec les différents régimes alimentaires, avec ou sans traitement par anticorps anti-PD1. Nous avons montré que le granzyme B était produit après traitement anti-PD1 uniquement dans les tumeurs des souris nourries avec le régime enrichi en indole-3-carbinol [6].
L’absence des ligands alimentaires de AhR nuit à la réactivation des lymphocytes T CD8+ et à l’infiltration de la tumeur par les lymphocytes NK après un traitement anti-PD1
La production de granzyme B étant associée à la présence des lymphocytes T CD8+ et des lymphocytes NK dans la tumeur, nous avons quantifié ces cellules par cytométrie en flux. Nous avons constaté que le traitement anti-PD1 a considérablement augmenté le nombre de lymphocytes NK (mais pas celui des lymphocytes T CD8+) uniquement dans les tumeurs des souris nourries avec le régime enrichi en indole-3-carbinol. Le traitement anti-PD-1 étant connu pour favoriser la réactivation et l’expansion des lymphocytes T CD8+, nous avons ensuite analysé la prolifération de ces lymphocytes après ce traitement, et constaté une augmentation de la prolifération uniquement dans les tumeurs des souris nourries avec le régime enrichi en indole-3-carbinol [6]. Ces résultats indiquent que l’absence des ligands alimentaires de AhR nuit aux réponses immunitaires précoces à l’immunothérapie par anticorps anti-PD1, en altérant la réactivation des lymphocytes T CD8+ et l’infiltration des lymphocytes NK dans la tumeur.
L’activation de AhR dans les lymphocytes T est nécessaire pour une efficacité optimale du traitement anti-PD1
Nous avons ensuite cherché à identifier la cible des ligands alimentaires de AhR parmi les cellules immunitaires infiltrant la tumeur. Afin de déterminer quelles populations de cellules immunitaires produisent AhR, nous avons utilisé une lignée de souris rapportrices présentant une fluorescence de la protéine TdTomato dans les cellules produisant AhR. Outre les monocytes, macrophages et lymphocytes NK, nous avons montré que AhR est produit par les lymphocytes T CD8+ spécifiquement dans les tumeurs, et non dans les autres organes. Pour déterminer l’importance relative des productions de AhR par les lymphocytes NK, les macrophages, et les lymphocytes T pour l’efficacité du traitement anti-PD1, nous avons utilisé plusieurs modèles de souris déficientes pour AhR uniquement dans chacun de ces trois types cellulaires. Nous avons ainsi montré la nécessité de la production de AhR par les lymphocytes T, mais pas de la production par les macrophages ou les lymphocytes NK [6]. Nous en avons conclu que l’effet des ligands alimentaires de AhR sur l’efficacité de l’immunothérapie par anticorps anti-PD1 repose principalement sur l’activation de AhR dans les lymphocytes T.
AhR favorise la réactivation des lymphocytes T CD8+ induite par le traitement anti-PD1
Afin de déterminer quels gènes sont contrôlés par AhR dans les lymphocytes T CD8+ lors du traitement anti-PD1, nous avons utilisé des souris déficientes ou non pour AhR dans les lymphocytes T. Nous avons isolé les lymphocytes T CD8+ de la tumeur en séparant les cellules progénitrices, décrites comme celles qui sont principalement ciblées par les anti-corps anti-PD1, et les cellules effectrices, puis réalisé une analyse transcriptomique de ces cellules. Nous avons ainsi montré que la présence de AhR est nécessaire dans les cellules progénitrices des lymphocytes T CD8+ pour permettre l’expression de gènes impliqués dans la réactivation de leur métabolisme mitochondrial et leur prolifération. Dans les lymphocytes T CD8+ effecteurs, l’activation de AhR par les ligands alimentaires est nécessaire à la production d’un ensemble de molécules effectrices incluant celles de la réponse à l’interféron, des molécules de surface telles que ICOS (inducible T-cell costimulator) et GITR (glucocorticoid-induced tumor necrosis factor receptor-related protein), ainsi que les chimiokines CCL3 et CCL4, connues pour attirer les lymphocytes NK [6]. Nous en avons conclu que l’activation de AhR dans les lymphocytes T CD8+ est essentielle pour permettre, en réponse à l’immunothérapie par anticorps anti-PD1, la récupération optimale des propriétés métaboliques et effectrices de ces lymphocytes (Figure 1).
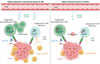 |
Figure 1 Le facteur de transcription AhR favorise la réactivation des lymphocytes T CD8+ induite par l’immunothérapie anti-PD1. Lors du traitement par des anticorps anti-PD1, l’activation de AhR par les ligands alimentaires est nécessaire à l’expression de gènes impliqués dans la réactivation et la prolifération des cellules progénitrices des lymphocytes T CD8+, et à la production de molécules effectrices par les lymphocytes T CD8+ effecteurs, telles que les chimiokines CCL3 et CCL4 qui peuvent attirer les lymphocytes NK (natural killer). Figure réalisée avec BioRender. |
Perspectives
Nos résultats expérimentaux indiquent que la présence d’indoles dans le régime alimentaire pourrait augmenter l’efficacité de l’immunothérapie par blocage des points de contrôle immunitaire chez des personnes atteintes d’un cancer. Ils pourraient également avoir des implications pour l’immunothérapie par lymphocytes CAR-T (chimeric antigenic receptor), car le métabolisme de ces cellules est un paramètre essentiel pour l’efficacité de ce traitement. Au-delà du cancer, le blocage de PD1 a montré un potentiel thérapeutique contre les infections chroniques (telles que l’hépatite B, l’infection par le virus de l’immunodéficience humaine, ou le paludisme au stade sanguin) et les septicémies. Les résultats obtenus chez la souris ouvrent la voie à de futurs essais cliniques.
Liens d’intérêt
Les auteures déclarent ne pas avoir de lien d’intérêt.
Références
- Pitt JM, Vétizou M, Daillère R, et al. Resistance mechanisms to immune-checkpoint blockade in cancer: Tumor-intrinsic and -extrinsic factors. Immunity 2016 ; 44 : 1255–69. [Google Scholar]
- Helmink BA, Khan MAW, Hermann A, et al. The microbiome, cancer, and cancer therapy. Nat Med 2019 ; 25 : 377–88. [Google Scholar]
- Magrì A, Germano G, Lorenzato A, et al. High-dose vitamin C enhances cancer immunotherapy. Sci Transl Med 2020 ; 12 : eaay8707. [Google Scholar]
- Yuan X, Duan Y, Xiao Y, et al. Vitamin E enhances cancer immunotherapy by reinvigorating dendritic cells via targeting checkpoint SHP1. Cancer Discov 2022 ; 12 : 1742. [Google Scholar]
- Ferrere G, Tidjani Alou MT, Liu P, et al. Ketogenic diet and ketone bodies enhance the anticancer effects of PD-1 blockade. JCI Insight 2021 ; 6 : e145207. [Google Scholar]
- De Juan A, Coillard A, Cros A, et al. Physiological activation of aryl hydrocarbon receptor by food-derived ligands is essential for the efficacy of anti-PD1 therapy. Nat Commun 2025 ; 16 : 10598. [Google Scholar]
© 2026 médecine/sciences – Inserm
 Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l’utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l’utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Liste des figures
|
Figure 1 Le facteur de transcription AhR favorise la réactivation des lymphocytes T CD8+ induite par l’immunothérapie anti-PD1. Lors du traitement par des anticorps anti-PD1, l’activation de AhR par les ligands alimentaires est nécessaire à l’expression de gènes impliqués dans la réactivation et la prolifération des cellules progénitrices des lymphocytes T CD8+, et à la production de molécules effectrices par les lymphocytes T CD8+ effecteurs, telles que les chimiokines CCL3 et CCL4 qui peuvent attirer les lymphocytes NK (natural killer). Figure réalisée avec BioRender. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.