")
")
| Issue |
Med Sci (Paris)
Volume 42, Number 5, Mai 2026
Réfléchir le vivant
|
|
|---|---|---|
| Page(s) | 510 - 513 | |
| Section | Forum | |
| DOI | https://doi.org/10.1051/medsci/2026070 | |
| Published online | 29 mai 2026 | |
Non, le blob n’a pas 720 sexes !
No, Physarum blobs do not have 720 sexes!
Professeur du Muséum national d’histoire naturelle, Institut de systématique, évolution, biodiversité, membre de l’Institut universitaire de France
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.

Vignette (© Marc-André Selosse).
La littérature à propos du blob1 (Physarum polycephalum ; Figure 1) s’exalte souvent, entre autres propriétés inattendues, des 720 sexes de cet organisme unicellulaire [1], qui s’ajoute à ses autres bizarreries… Mais comment peut-on avoir plus de deux sexes ?
 |
Figure 1 Appareil végétatif du blob (Physarum polycephalum), une amibe géante. Le Bernemi, CC BY-SA 4.0 via Wikimedia Commons. |
Du sexe au blob
Revenons aux définitions de base. Au sens strict chez les eucaryotes, la sexualité est la fusion de deux cellules à n chromosomes, appelées gamètes, qui engendre la première cellule avec 2n chromosomes de la génération suivante. Les gamètes ont un seul jeu de chromosomes : haploïdes, ils donnent une cellule diploïde lors de la fécondation. Chez le blob, les spores qui dispersent l’espèce germent en petites amibes2 unicellulaires. Dans certaines conditions, deux de ces amibes se rencontrent et se transforment en gamètes : elles opèrent une fusion et engendrent une cellule diploïde qui, elle, ne se divisera plus. Elle grandira, son noyau se divisera à l’intérieur d’elle-même et elle engendrera le blob, une amibe géante à noyaux multiples3.
Il y a toujours deux sexes (au sens de partenaires) dans un évènement sexuel. Soit leurs gamètes sont identiques (même taille, rôle symétrique comme chez le blob) et les deux sexes sont indifférenciables. Soit l’un des gamètes est plus petit que l’autre et on peut alors distinguer les sexes mâle (qui produit le plus petit gamète et investit le moins) et femelle (qui investit le plus, avec le plus gros gamète4). Dans ce second cas, celui de l’humain, on distingue donc des individus de sexe mâle ou femelle (au sens de sexe différencié). Les organismes hermaphrodites (comme les escargots ou les fleurs), quant à eux, peuvent être alternativement mâle ou femelle selon les fécondations.
Donc deux sexes pour une fécondation, parfois distinguable en deux catégories, mâle et femelle… Alors, comment avoir 720 sexes ? Laissons le blob de côté et examinons d’abord comment s’apparient les gamètes. En effet, l’intérêt de la sexualité est le brassage génétique. Plus les partenaires sont différents, plus la combinaison génétique produite est nouvelle et différente des parents, ce qui peut fournir un avantage adaptatif face aux changements du milieu. Plus les partenaires sont semblables, en revanche, plus le risque de consanguinité est grand [2] (→).
(→) Voir m/s n° 3, 2024, page 288
De façon logique, en plus des sexes mâle et femelle, quand ils sont séparés, des dispositifs génétiques ont émergé qui évitent l’autofécondation (chez les hermaphrodites), voire la fécondation entre individus proches. Ce sont les incompatibilités sexuelles.
Du sexe des champignons…
Commençons par la levure de boulanger, Saccharomyces cerevisiae. La fécondation implique deux cellules haploïdes identiques qui, en présence d’une de l’autre, deviennent des gamètes : elles forment deux expansions en vis-à-vis par où elles fusionnent. Ni mâle, ni femelle, pas de différence entre les sexes, mais… un gène à deux allèles, a et α, contraint les levures a à féconder les α seulement, et vice-versa. S’agit-il d’une différenciation en sexes mâle et femelle cachée ? Non, les rôles et les contributions sont symétriques, c’est bel et bien une incompatibilité sexuelle.
En effet, chez d’autres champignons du groupe des ascomycètes, ce mécanisme (un gène d’incompatibilité à deux allèles, souvent notés + et -) se superpose à l’existence de différenciations sexuelles mâle et femelle [3]. Par exemple, chez Neurospora crassa, les mycéliums forment des petites cellules fécondantes, les spermaties (des gamètes mâles), libérées dans le milieu, et des renflements, ou ascogones (les gamètes femelles), qui sont fécondés par les spermaties puis nourrissent la cellule formée après fécondation (Figure 2).
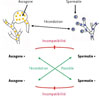 |
Figure 2 Différenciations sexuelles du mycélium de l’ascomycète Neurospora crassa : l’ascogone (noyaux en rouge) est la partie femelle, fécondable par les spermaties, des spermatozoïdes sans motilité propre (noyaux en bleu ; parfois inexactement appelés conidies). Il existe deux types de mycélium, + et -, résultants d’un gène d’incompatibilité à deux allèles, qui empêche les fécondations entre structures du même type d’incompatibilité, issues du même mycélium ou non. |
Bien que chaque individu soit hermaphrodite, et forme donc spermaties et ascogones, la fécondation ne s’opère qu’entre individus + et – (Figure 2). Spermaties et ascogones de même type sont sexuellement incompatibles entre eux. Chez les ascomycètes, le gène responsable de cette incompatibilité code une protéine régulatrice de la transcription dotée d’un domaine de liaison à l’ADN, par exemple un homéodomaine5. Cette protéine régulatrice existe sous deux formes alternatives, qui ont des effets multiples. Elle permet aux individus de se comporter en type + ou en type –, avant la fécondation, et de jouer un rôle complémentaire à l’autre type lors de la fécondation. Après celleci, dans les individus devenus +/–, l’hétérozygotie obtenue bloque les mécanismes menant à la fécondation, mais permet la méiose, qui permet le retour à l’état haploïde si les conditions s’y prêtent. Organisateur du cycle biologique, ce gène limite aussi l’autofécondation, qui entrainerait une forme extrême de consanguinité : c’est un gène d’incompatibilité.
Groupe voisin, les champignons Basidiomycètes ont quant à eux deux gènes d’incompatibilité, ou plutôt deux locus, nommés A et B, comprenant chacun plusieurs gènes. Ces gènes sont multialléliques, nous y reviendrons. Le locus A code des facteurs de transcription à homéodomaine, organisant le cycle de vie du champignon de façon homologue au gène d’incompatibilité des Ascomycètes. Le locus B code un système de phéromones6 requis pour la fécondation. Il comporte plusieurs gènes liés ensemble : les uns codent des récepteurs reconnaissant des peptides secrétés, tandis que les autres codent ces peptides secrétés, qui agissent notamment comme signal pour organiser la fécondation entre partenaires (ce sont des phéromones). Mais il y a une particularité : les récepteurs dérivés d’un allèle donné du locus B reconnaissent les phéromones produites par tous les autres allèles de B, mais non pas celles produites par ce locus lui-même. Ainsi, aucun risque d’interférence d’un haploïde isolé avec lui-même. La fécondation ne réussit que si les allèles des locus A et B des cellules partenaires sont différents. De nouveau, outre de nombreuses fonctions, ce dispositif évite l’autofécondation, mais avec un avantage cette fois : chacun de deux locus A et B ont des dizaines d’allèles. Or, c’est la combinaison des allèles des deux locus, chacun très varié, qui constitue le type d’incompatibilité. Il y aurait donc, pour des espèces comme Schizophyllum commune ou le coprin Coprinellus cinereus, plus de 20 000 types d’incompatibilité différents !
Ce dispositif exclut aussi les individus apparentés et donc la consanguinité : dans une telle diversité de types d’incompatibilité, les individus identiques aux deux locus sont souvent très proches génétiquement. Cependant, la plupart des rencontres sont compatibles… alors qu’avec seulement deux allèles, la moitié de la population est incompatible, y compris au passage de nombreux individus non apparentés. Ce système d’incompatibilité multiallélique limite donc la consanguinité tout en suscitant moins d’interfertilités inutiles.
… au sexe des fleurs
On retrouve un gène d’incompatibilité multiallélique chez les plantes à fleurs, les Angiospermes. La plupart du temps, elles sont hermaphrodites, à la façon des ascomycètes évoqués plus haut : les fleurs présentent du pollen porté par les étamines (partie mâle) et des ovules au centre de la fleur, dans l’ovaire surmonté par le pistil7 (partie femelle). L’autofécondation est évitée grâce à un locus, nommé S (pour self-incompatibility), qui entraine un rejet quand le pollen et le pistil portent au moins un allèle identique : seul un grain de pollen ne présentant pas les allèles S d’un pistil peut y germer [4]. Parfois c’est l’allèle porté par le grain de pollen lui-même qui s’exprime : celui-ci est haploïde, car issu d’une méiose, et ne porte qu’un des deux allèles de la plante mère, sur laquelle il est donc incapable de germer (on parle d’incompatibilité gamétophytique, et le rejet a souvent lieu après la germination du pollen, quand son génome commence à s’exprimer). Dans d’autres espèces, la reconnaissance implique des protéines déposées par la plante-mère à la surface du grain de pollen, issues donc des deux allèles de la plante-mère (c’est l’incompatibilité sporophytique, où le rejet est souvent plus précoce). Il existe des dizaines d’allèles du locus S ; de plus, comme le pistil exprime toujours deux allèles, c’est le nombre de duos possibles qui détermine le nombre de types d’incompatibilité, soit presque le carré du nombre d’allèles !
Les Angiospermes présentent donc comme les Basidiomycètes un système d’incompatibilité multiallélique qui, tout en évitant l’autofécondation et les individus apparentés, minimise les rejets inutiles. Ces systèmes multialléliques maintiennent d’ailleurs leur diversité d’allèles au cours du temps. En effet, quand un allèle devient rare, il est rarement confronté à lui-même par hasard : il a donc souvent une descendance et est ainsi sélectionné. L’inverse se produit quand un allèle devient très fréquent, et donc plus souvent confronté avec lui-même, y compris par hasard : plus souvent privé de descendance, il est donc contre-sélectionné et fait place aux allèles rares. Ainsi, les allèles ne peuvent devenir trop fréquents, et, chaque fois qu’ils deviennent plus rares, ils sont sauvés… par leur rareté : ce mécanisme de sélection balancée maintient la diversité allélique.
Du blob aux humains
Eh bien, le blob… a des systèmes d’incompatibilité sexuelle aussi, gouvernés par un gène multiallélique qui aurait au moins 720 allèles [1], c’est-à-dire qu’il a 720 types d’incompatibilité puisque les gamètes sont haploïdes (ils portent un allèle chacun). Il a deux partenaires sexuels à chaque fécondation (comme tout le monde !), mais pas de sexe mâle ou femelle différencié. Présenter le blob comme ayant 720 sexes voudrait dire que les fleurs en ont des dizaines, sinon des centaines. Il s’agit en fait de types d’incompatibilité : et en la matière, certains champignons basidiomycètes surpassent le blob d’un facteur 10 : on ne dit pas pour autant qu’ils ont des milliers de sexes ! L’universalité des incompatibilités va plus loin. Chez les Mammifères, comme souvent dans le monde animal, les sexes sont séparés, ce qui évite l’autofécondation.
Mais un dispositif supplémentaire semble éviter la consanguinité. Le complexe majeur d’histocompatibilité (CMH), formé de deux locus, est surtout connu pour ses rôles majeurs dans le système immunitaire : il existe des milliers d’allèles de chaque locus dans les populations humaines. Les compatibilités de greffe, qui requièrent une identité de CMH, montrent que les proches parents sont souvent les seuls à posséder des formes identiques. Or, certains mammifères s’accouplent de préférence avec des partenaires au CMH plus divergent d’eux que le hasard ne le laisserait attendre. Pour les souris, où ce phénomène a été d’abord été reconnu, des sécrétions lacrymales contiendraient des peptides (issus du CMH) impliqués dans la reconnaissance et conduisant à l’évitement des proches parents ; elles sont perçues dans leur organe voméronasal8 [5]. Chez l’humain, un biais d’appariement significatif est décrit dans certaines populations, notamment occidentales, mais pas toutes [6] : nous éviterions en moyenne les individus de CMH proche ! Les mécanismes de cet évitement restent à clarifier (en l’absence d’organe voméronasal chez l’humain) et ils semblent modulés par des traits culturels [7]. Si le CMH joue un rôle d’incompatibilité chez les vertébrés, en plus de ses autres fonctions, alors cela contribue aussi à sélectionner son état multiallélique : car, tout en évitant la consanguinité, le multiallélisme minimise le risque d’interfertilités inutiles, à la façon des locus S, A et B…
Ces histoires sexuelles ne retranchent rien à la fascination du blob, bien au contraire, mais elles montrent comment les experts des différents groupes vivants sont si spécialisés qu’ils ignorent souvent l’universalité d’un mécanisme… aussi banal qu’une fleur. Hélas, cette spécialisation prévaut dès les premières années universitaires, pourtant généralistes, où elle masque la généralité de certains mécanismes biologiques : nous avons vu de belles évolutions convergentes, trop peu enseignées comme telles ! Tout cela trompe sur les mécanismes du sexe, assez pour nous laisser fantasmer d’obscures orgies multisexuelles chez les blobs !
Liens d’intérêt
L’auteur déclare ne pas avoir de lien d’intérêt.
Références
- Moriyama Y, Kawano S. Maternal inheritance of mitochondria: multipolarity, multiallelism and hierarchical transmission of mitochondrial DNA in the true slime mold Physarum polycephalum. J Plant Res 2010 ; 123 : 139–48. [Google Scholar]
- Selosse MA. Grandeur et misère de la diploïdie. Med Sci (Paris) 2024 ; 40 : 288–9. [Google Scholar]
- Sun S, Coelho MA, David-Palma M, et al. Fungal sexual reproduction and mating-type loci. Curr Biol 2025 ; 35 : R496–503. [Google Scholar]
- Wheeler MJ, Franklin-Tong VE, Franklin FCH. The molecular and genetic basis of pollen-pistil interactions. New Phytologist 2001 ; 151 : 565–84. [Google Scholar]
- Yan J, Zhang B, Dunn DW, Li B, Zhang P. Mate choice and the major histocompatibility complex: a review. Biol J Linnean Soc 2024 ; 143 : blae100. [Google Scholar]
- Chaix R, Cao C, Donnelly P. Is mate choice in humans MHC-dependent? PLoS Genet 2008 ; 4 : e1000184. [Google Scholar]
- Selosse MA. Nature et préjugés. Convier l’humanité dans l’histoire naturelle. Arles : Actes Sud 2024 : 448 p. [Google Scholar]
Le nom « blob » vient de l’anglais et signifie littéralement « masse informe » ou « tache gélatineuse ».
Cellule sans paroi, capable de déformation pour se déplacer et absorber des aliments par phagocytose.
Cette grande amibe à multiples noyaux est aussi appelée plasmode.
Cette définition est simpliste pour les pluricellulaires, plantes ou animaux, où les coûts peuvent aussi provenir de l’entretien de la génération suivante. Chez l’humain, le coût énergétique pour la femme est peu important au niveau de l’ovule (même s’il est plus gros que le spermatozoïde), mais bien plus important lors de la grossesse et de l’allaitement.
Type de séquence d’une soixantaine d’acides aminés qui peut se fixer sur l’ADN au niveau d’une région régulatrice de l’expression des gènes, ainsi nommée car les premières furent identifiées sur les gènes homéotiques régulateurs du développement chez la Drosophile.
Message chimique produit par un organisme qui induit une réaction chez un autre organisme de la même espèce. Chez les ascomycètes, il y a aussi des phéromones peptidiques, mais chaque individu haploïde possède les gènes des deux types de phéromones, + et - : selon son type sexuel, il n’exprime que celui correspondant à son type (et les diploïdes ne produisent plus de phéromones).
Le pistil reçoit les grains de pollen et permet leur germination en un tube qui injecte ensuite les spermatozoïdes dans l’ovule.
Cet organe, situé dans le nez de certains mammifères, détecte des signaux chimiques et notamment des phéromones.
© 2026 médecine/sciences – Inserm
 Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l’utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l’utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Liste des figures
|
Figure 1 Appareil végétatif du blob (Physarum polycephalum), une amibe géante. Le Bernemi, CC BY-SA 4.0 via Wikimedia Commons. |
| Dans le texte | |
|
Figure 2 Différenciations sexuelles du mycélium de l’ascomycète Neurospora crassa : l’ascogone (noyaux en rouge) est la partie femelle, fécondable par les spermaties, des spermatozoïdes sans motilité propre (noyaux en bleu ; parfois inexactement appelés conidies). Il existe deux types de mycélium, + et -, résultants d’un gène d’incompatibilité à deux allèles, qui empêche les fécondations entre structures du même type d’incompatibilité, issues du même mycélium ou non. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.