")
")
| Issue |
Med Sci (Paris)
Volume 33, Number 10, Octobre 2017
|
|
|---|---|---|
| Page(s) | 825 - 828 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/20173310005 | |
| Published online | 10 octobre 2017 | |
Clostridium difficile : la belle et la bête
Clostridium difficile: the beauty and the beast
Center for Inflammation Immunity and Infection, Institute for Biomedical Sciences, Georgia State University, 100 Piedmont Ave SE, 30303 Atlanta, Georgie, États-Unis
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Une étude récente, publiée dans la revue Gut, a mis en évidence une nouvelle souche de Clostridium difficile non pathogène, nommée C. difficile LEM1, capable de protéger contre une infection à C. difficile normalement létale. Sa découverte dans le microbiote intestinal suggère l’intérêt bénéfique de cette bactérie pour le traitement des infections à C. difficile.
Épidémiologie et physiopathologie des infections à Clostridium difficile
Clostridium difficile est un bacille à Gram positif, anaérobie, sporulé et responsable de 15 à 25 % des cas de diarrhées nosocomiales après antibiothérapie. C. difficile représente entre 90 et 95 % des cas de colites pseudomembraneuses acquises le plus souvent en milieu hospitalier [1] (→).
(→) Voir la Synthèse de C. Eckert et F. Barbut, m/s n° 2, février 2010, page 153
En Europe, en moyenne 7 cas d’infections à C. difficile (ICD) sont recensés pour 10 000 hospitalisations, ce qui représente un coût annuel qui pourrait atteindre 3 milliards d’euros [2, 3]. L’incidence est similaire aux États-Unis où environ 500 000 nouveaux cas d’ICD sont recensés chaque année. En 2011, 29 000 décès ont été attribués à cette bactérie, représentant un coût annuel de 4,8 millions de dollars [4, 5]. Les ICD surviennent le plus souvent à la suite de l’administration d’antibiotiques qui participent à la dysbiose responsable du dysfonctionnement du microbiote intestinal, et notamment à la rupture de sa fonction de barrière, permettant l’implantation et la multiplication de C. difficile au niveau du tractus gastro-intestinal.
Le processus infectieux commence par une étape de colonisation du tube digestif de l’hôte à la suite de l’ingestion de spores bactériennes qui germent sous l’action des sels biliaires [6]. Les spores sont très stables et sont fréquemment rencontrées en milieux hospitaliers, le diagnostic des personnes infectées étant réalisé après le début de la libération de ces spores par les patients. Environ 50 % des infections résulteraient de souches qui ne sont pas directement liées à un cas antérieur d’infection par C. difficile, indiquant que, pour de nombreux cas, l’origine reste encore incertaine. À la suite de la colonisation de la muqueuse intestinale, des lésions apparaissent, ayant pour origine la production par la bactérie de deux toxines, TcdA (toxine C. difficile A) et TcdB [7].
Clostridium difficile et le microbiote intestinal
Comme l’homme, les rongeurs sont habituellement résistants à l’infection par C. difficile. Buffie et ses collaborateurs ont en effet montré qu’une bactérie présente dans le microbiote intestinal, Clostridium scindens, contribue à la résistance contre l’infection par C. difficile par la biosynthèse d’acides biliaires secondaires (Figure 1) [6]. Les animaux peuvent néanmoins devenir sensibles à C. difficile après un traitement antibiotique qui provoque une dysbiose rompant ainsi l’effet de barrière du microbiote, ou s’ils sont dépourvus de microbiote (animaux axéniques) [8, 9]. Nous avons rendu des souris susceptibles à l’infection par C. difficile par un traitement antibiotique (un mélange de 5 antibiotiques administrés dans l’eau de boisson ad libitum simultanément avec une injection intra-péritonéale d’un autre antibiotique) avant infection par des spores de la souche de référence C. difficile VPI 10463 (souche virulente ATCC 43255, produisant les toxines TcdA et TcdB). La présence de C. difficile dans les fèces des souris infectées a été analysée par la quantification du gène tcdA, codant la toxine TcdA, par PCR (polymerase chain reaction) quantitative : cette méthode fondée sur l’extraction d’ADN à partir de matériel fécal permet de s’affranchir de l’utilisation des techniques de cultures bactériennes anaérobies classiques [10].
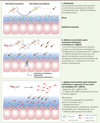 |
Figure 1. Rôle du microbiote dans la défense contre Clostridium difficile. A. Lorsque le microbiote est intact, certaines bactéries convertissent les sels biliaires primaires en sels biliaires secondaires s’opposant ainsi à la multiplication et à la colonisation par C. difficile. B. Lorsque le microbiote est perturbé suite à un traitement antibiotique, les sels biliaires primaires ne sont plus convertis en sels biliaires secondaires et ne peuvent plus exercer leur action inhibitrice sur C. difficile. C. Lorsque le microbiote est perturbé par un traitement antibiotique, la souche non pathogène C. difficile LEM1 apparaît et empêche la colonisation par la souche pathogène C. difficile VPI 10463. |
Identification et séquençage d’une nouvelle souche de Clostridium difficile murine protégeant contre les infections par une souche pathogène
Après le traitement par antibiotiques, nous avons observé un pourcentage important de souris non infectées par C. difficile VPI 10463 qui présentait néanmoins des niveaux importants du gène tcdA dans leurs fèces, sans pour autant développer des signes cliniques de pathologies intestinales. Le transfert du microbiote fécal de ces souris conventionnelles traitées par antibiothérapie et détectées positives pour tcdA à des souris axéniques n’induit pas de signes cliniques chez les souris receveuses, indiquant que la souche de C. difficile présente chez les souris conventionnelles n’est pas virulente. Ces résultats montrent donc que l’antibiothérapie induit des modifications du microbiote intestinal qui permettent l’expansion d’une souche endogène de C. difficile qui, bien que produisant des toxines, reste avirulente [10].
Une analyse rétrospective du microbiote intestinal des souris conventionelles exposées au traitement antibiotique a permis de mettre en évidence des altérations majeures de la composition et de la diversité des populations microbiennes, à l’origine de susceptibilités différentes à C. difficile et définissant deux catégories d’animaux : des animaux dits « résistants » et des animaux dits « sensibles ». En effet, les souris considérées comme résistantes présentaient un microbiote constitué d’une faible abondance d’une souche endogène de C. difficile, nommée LEM1, capable de produire des toxines mais n’induisant pas de signes cliniques. À l’opposé, chez les animaux sensibles, qui succombaient en quelques jours à une infection par la souche virulente C. difficile VPI 10463, la souche LEM1 n’était pas détectée.
La souche murine de C. difficile LEM1 a été isolée du microbiote des animaux « résistants » par des techniques de bactériologie classique (isolement sur milieu gélosé sélectif suivi d’une culture en chambre anaérobie), et a été caractérisée par une amplification PCR des ARN ribosomaux (ou ribotypage : PCR ribotyping, en anglais). L’identification du profil obtenu par ribotypage et sa comparaison aux profils des souches de références ont révélé que la souche LEM1 n’avait pas été identifiée à ce jour. L’utilisation d’une technique très sensible de PCR nichée1 (nested-PCR en anglais) a montré que 40 % des souris résistantes possédaient, dans leur microbiote, la souche C. difficile LEM1 avant même le traitement antibiotique alors qu’aucune des souris « susceptibles » ne la présentait, suggérant que certains animaux hébergeaient naturellement cette souche dans leur microbiote et ce avant même l’exposition au traitement antibiotique. Le génome de la souche C. difficile LEM1 a été séquencé et a été comparé à celui de la souche de référence C. difficile VPI 10463 [11]. L’alignement des deux génomes a permis de mettre en évidence de nombreuses translocations et des réarrangements génomiques avec, cependant, un degré global de similarité élevé (95 %) entre les deux souches.
Cette étude montre que l’administration exogène de la souche C. difficile LEM1 n’induit pas de signes pathologiques dans le modèle de souris conventionelles recevant une antibiothérapie, ni chez les souris axéniques (souris pourtant sujettes à développer des signes cliniques fatals en quelques heures après une infection par la souche pathogène C. difficile VPI 10463). La faible virulence de C. difficile LEM1, même lorsqu’elle est administrée à fortes doses, peut être expliquée par sa faible capacité à produire les toxines par rapport à C. difficile VPI 10463. Nos résultats ont également démontré que l’administration préventive, ou concomitante, de la souche C difficile LEM1 avec C. difficile VPI 10463, permet de protéger les animaux contre une infection qui est normalement létale. La souche C. difficile LEM1 est en effet capable de diminuer la colonisation intestinale par la souche VPI 10463, et donc les signes pathologiques qui sont associés à l’infection, chez des souris conventionnelles et des souris axéniques, probablement par un mécanisme de compétition vis-à-vis d’une niche écologique commune [10] (Figure 1). La capacité de la souche C. difficile LEM1 à protéger contre une souche virulente corrobore les observations démontrant que des souches non-toxinogènes peuvent prévenir la mortalité induite par C. difficile chez les hamsters [12] et réduire le taux de récidive chez l’homme [13].
Conclusions et perspectives
Les résultats que nous avons obtenus mettent en évidence la présence, au niveau du microbiote intestinal murin, d’une bactérie à potentiel bénéfique. Ils suggèrent son possible usage thérapeutique pour le traitement des ICD. Les implications de cette étude sont nombreuses mais la compréhension des mécanismes de protection de la souche C. difficile LEM1 vis-à-vis de C. difficile VPI 10463 reste à élucider. La présence de bactériophages et/ou la sécrétion de molécules inhibitrices produites par la souche C. difficile LEM1 et pouvant agir sur C. difficile VPI 10463 sont des pistes envisagées. L’extrapolation de ces résultats chez l’homme suggèrent que certaines ICD asymptomatiques, qui se manifestent dans les hôpitaux, pourraient être en fait dues à des souches de C. difficile présentes à de très faibles niveaux (voire indétectables) dans le microbiote intestinal et non à l’acquisition de souches de C. difficile présentes dans l’environnement hospitalier.
Le traitement antibiotique induit de nombreuses perturbations du microbiote. Certaines des espèces bactériennes sont relativement résistantes. Nous recherchons, par l’analyse des données de séquençage et par des techniques de culture anaérobie, à définir des taxons2 bactériens dont l’élimination, par le traitement antibiotique, serait corrélée avec la prolifération de la souche C. difficile LEM1 endogène et pourraient représenter de nouvelles voies thérapeutiques pour le traitement des ICD.
Liens d’intérêt
Les auteurs déclarent n’avoir aucun lien d’intérêt concernant les données publiées dans cet article.
Remerciements
Je remercie Benoit Chassaing (Institute for biomedical sciences, Atlanta, GA, États-Unis) pour la relecture critique du manuscrit.
La technique de nested-PCR (PCR nichée) se pratique comme la PCR (polymerase chain reaction) avec un couple d’amorces externes pour quelques cycles d’amplification. Ensuite, un excès d’une amorce interne est ajouté avant de prolonger l’amplification.
Un taxon correspond à une entité d’êtres vivants regroupés parce qu’ils possèdent des caractères en communs du fait de leur parenté.
Références
- Eckert C, Barbut F. Infections associées à Clostridium difficile. Med Sci (Paris) 2010 ; 26 : 153–158. [CrossRef] [EDP Sciences] [PubMed] [Google Scholar]
- Bouza E. Consequences of Clostridium difficile infection: understanding the healthcare burden. Clin Microbiol Infect 2012 ; 18 (suppl 6) : 5–12. [CrossRef] [PubMed] [Google Scholar]
- Davies KA, Longshaw CM, Davis GL, et al. Underdiagnosis of Clostridium difficile across Europe: the European, multicentre, prospective, biannual, point-prevalence study of Clostridium difficile infection in hospitalised patients with diarrhoea (EUCLID). Lancet Infect Dis 2014 ; 14 : 1208–1219. [CrossRef] [PubMed] [Google Scholar]
- Lessa FC, Mu Y, Bamberg WM, et al. Burden of Clostridium difficile infection in the United States. N Engl J Med 2015 ; 372 : 825–834. [CrossRef] [PubMed] [Google Scholar]
- Martin JS, Monaghan TM, Wilcox MH. Clostridium difficile infection: epidemiology, diagnosis and understanding transmission. Nat Rev Gastroenterol Hepatol 2016 ; 13 : 206–216. [CrossRef] [PubMed] [Google Scholar]
- Buffie CG, Bucci V, Stein RR, et al. Precision microbiome reconstitution restores bile acid mediated resistance to Clostridium difficile. Nature 2015 ; 517 : 205–208. [CrossRef] [PubMed] [Google Scholar]
- Rupnik M, Wilcox MH, Gerding DN. Clostridium difficile infection: new developments in epidemiology and pathogenesis. Nat Rev Microbiol 2009 ; 7 : 526–536. [CrossRef] [PubMed] [Google Scholar]
- Chen X, Katchar K, Goldsmith JD, et al. A mouse model of Clostridium difficile-associated disease. Gastroenterology 2008 ; 135 : 1984–1992. [CrossRef] [PubMed] [Google Scholar]
- Reeves AE, Koenigsknecht MJ, Bergin IL, Young VB. Suppression of Clostridium difficile in the gastrointestinal tracts of germfree mice inoculated with a murine isolate from the family Lachnospiraceae. Infect Immun 2012 ; 80 : 3786–3794. [CrossRef] [PubMed] [Google Scholar]
- Etienne-Mesmin L, Chassaing B, Adekunle O, et al. Toxin-positive Clostridium difficile latently infect mouse colonies and protect against highly-pathogenic C. difficile. Gut 2017; pii : gutjnl-2016-313510. [Google Scholar]
- Etienne-Mesmin L, Chassaing B, Adekunle O, et al. Genome sequence of a toxin-positive Clostridium difficile strain isolated from murine feces. Genome Announc 2017; 5 : pii : e00088–17. [Google Scholar]
- Nagaro KJ, Phillips ST, Cheknis AK, et al. Nontoxigenic Clostridium difficile protects hamsters against challenge with historic and epidemic strains of toxigenic BI/NAP1/027 C. difficile. Antimicrob Agents Chemother 2013 ; 57 : 5266–5270. [CrossRef] [PubMed] [Google Scholar]
- Gerding DN, Meyer T, Lee C, et al. Administration of spores of nontoxigenic Clostridium difficile strain M3 for prevention of recurrent C. difficile infection: a randomized clinical trial. JAMA 2015 ; 313 : 1719–1727. [CrossRef] [PubMed] [Google Scholar]
© 2017 médecine/sciences – Inserm
Liste des figures
|
Figure 1. Rôle du microbiote dans la défense contre Clostridium difficile. A. Lorsque le microbiote est intact, certaines bactéries convertissent les sels biliaires primaires en sels biliaires secondaires s’opposant ainsi à la multiplication et à la colonisation par C. difficile. B. Lorsque le microbiote est perturbé suite à un traitement antibiotique, les sels biliaires primaires ne sont plus convertis en sels biliaires secondaires et ne peuvent plus exercer leur action inhibitrice sur C. difficile. C. Lorsque le microbiote est perturbé par un traitement antibiotique, la souche non pathogène C. difficile LEM1 apparaît et empêche la colonisation par la souche pathogène C. difficile VPI 10463. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.