")
")
| Issue |
Med Sci (Paris)
Volume 29, Number 5, Mai 2013
|
|
|---|---|---|
| Page(s) | 523 - 528 | |
| Section | M/S Revues | |
| DOI | https://doi.org/10.1051/medsci/2013295016 | |
| Published online | 28 mai 2013 | |
Du cannabis aux agonistes sélectifs du récepteur CB2
Des molécules aux nombreuses vertus thérapeutiques
From cannabis to selective CB2R agonists: molecules with numerous therapeutical virtues
1
Institut de chimie pharmaceutique Albert Lespagnol, université Lille Nord, EA 4481, IFR114, 3, rue du Pr Laguesse, BP 83, 59006
Lille, France
2
Unité de glycobiologie structurale et fonctionnelle, CNRS UMR 8576, université Lille 1, 59650
Villeneuve d’Ascq, France
* Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Résumé
Utilisée à l’origine en Asie pour traiter douleurs, spasmes, nausées et insomnies, la marijuana est le psychotrope le plus consommé au monde. L’intérêt du cannabis médical a été reconsidéré depuis peu, menant à de nombreuses recherches et à la commercialisation de médicaments. Les cannabinoïdes naturels et synthétiques exercent des effets bénéfiques dans de nombreuses maladies. Ils sont toutefois accompagnés d’effets indésirables psychiatriques et cognitifs présumés liés au récepteur CB1. Les recherches actuelles tendent à concevoir des molécules thérapeutiques agonistes CB2 sans effets secondaires centraux psychotropes.
Abstract
Originally used in Asia for the treatment of pain, spasms, nausea and insomnia, marijuana is the most consumed psychotropic drug worldwide. The interest of medical cannabis has been reconsidered recently, leading to many scientific researches and commercialization of these drugs. Natural and synthetic cannabinoids display beneficial antiemetic, anti-inflammatory and analgesic effects in numerous diseases, however accompanied with undesirable effects due to the CB1 receptor. Present researches focus on the design of therapeutical molecules targeting the CB2 receptors, and thus avoiding central side effects and therefore psychotropic effects caused by the CB1 receptor.
© 2013 médecine/sciences – Inserm
Vignette (Photo © Inserm - Frédérique Koulikoff).
Cannabis sativa L. : une plante aux effets psychotropes à fort potentiel thérapeutique
Le cannabis (Cannabis sativa L.) est une plante annuelle de la famille des Cannabaceae très souvent associée à ses propriétés psychotropes récréatives. Cependant, les propriétés thérapeutiques de cette drogue sont connues depuis l’antiquité. Le Δ9-tétrahydrocannabinol (THC), substance psychoactive majoritaire du cannabis, a été isolé en 1964 par Y. Gaoni et R. Mechoulam de l’Institut des sciences Weizmann en Israël [1].
De nombreuses vertus sont reconnues au cannabis et justifient son usage pour soulager les effets secondaires liés aux maladies ou aux traitements. Il possède des effets analgésiques, antispasmodiques (spasticité dans la sclérose en plaques, SEP), anticonvulsivants (épilepsie), anti-inflammatoires, antivomitifs (effets secondaires des chimiothérapies anticancéreuses), bronchodilatateurs (asthme), vasodilatateurs (glaucome), relaxants et soporifiques. Sa capacité à soulager les symptômes de maladies neurodégénératives (SEP, maladies de Huntington, Parkinson et Alzheimer, sclérose latérale amyotrophique) [2] et ses propriétés antiprolifératives sur des lignées de cellules tumorales issues de cancers pulmonaires, mammaires, coliques, thyroïdiens, d’hémopathies ou de glioblastomes ont été démontrées [3]. Mais le cannabis est surtout utilisé chez les patients en phase terminale pour soulager les douleurs chroniques. L’utilisation du cannabis médical est de plus en plus tolérée et même légalisée dans certains pays et délivrée sur ordonnance (certains États américains, australiens et européens) [4].
Le cannabis à usage médical existe sous plusieurs formes [5] : le Marinol®, THC de synthèse, et le Cesamet®, analogue synthétique du THC, sont prescrits pour traiter les nausées et vomissements liés à la chimiothérapie; le Sativex®, extrait de cannabis, est prescrit comme antidouleur chez les patients atteints de SEP et de cancer; le Cannador®, autre extrait de cannabis, est prescrit pour diminuer la raideur musculaire, les spasmes musculaires et les douleurs associés à la SEP, pour traiter la cachexie chez les patients cancéreux et pour soulager les douleurs postopératoires.
Le système endocannabinoïde
Le mécanisme d’action du cannabis n’a été identifié que récemment. Le THC agit essentiellement sur le système endocannabinoïde (SEC) qui module diverses fonctions physiologiques (fonctions motrices, mémoire, motivation, énergie, douleur et émotion) [6]. Le système endocannabinoïde inclut les endocannabinoïdes (EC), dérivés d’acides gras polyinsaturés, plusieurs enzymes et protéines impliquées dans la biosynthèse, le transport et la bio-inactivation de ces EC, ainsi que les récepteurs CB1 (CB1R) et CB2 (CB2R) qui lient les EC.
Les endocannabinoïdes, dont les principaux sont l’anandamide (AEA), agoniste partiel du récepteur CB1, et le 2-arachidonoylglycérol (2-AG), ligand des récepteurs CB1 et CB2, ne sont pas des neurotransmetteurs classiques : en effet, à la différence de ces derniers, les EC ne sont pas stockés dans des vésicules synaptiques; ils sont synthétisés et libérés à la demande en réponse à une stimulation, via l’hydrolyse de leurs précurseurs lipidiques membranaires. Libérés par le neurone postsynaptique, ils agissent de façon rétrograde en stimulant leurs récepteurs présynaptiques [7]. Les fonctions des EC sont présumées neuro- et immunomodulatrices [8]. Dans l’organisme, les EC sont hydrolysés très rapidement par des enzymes (trois sont connues à ce jour, dont la fatty acid amide hydolase, FAAH) et ont une durée de vie extrêmement courte [9]. Depuis quelques années, des inhibiteurs de la FAAH sont développés afin de bloquer le catabolisme rapide de l’AEA et prolonger sa durée d’action, offrant ainsi une nouvelle piste thérapeutique très prometteuse [10].
Le clonage des gènes codant pour les récepteurs CB1 en 1990 [11] et CB2 en 1993 [12] a permis d’exploiter les propriétés thérapeutiques non psychotropes des cannabinoïdes. En effet, les effets psychotropes (euphorie, vertiges, anxiété, sédation, etc.), ainsi que la dépendance au THC et autres composés addictogènes (nicotine, alcool, cocaïne), ont été essentiellement attribués au CB1R [13], alors que de nombreuses propriétés médicinales, en particulier celles liées à la modulation du système immunitaire, sont probablement associées au CB2R [14]. Le CB1R est abondamment exprimé dans le système nerveux central, et sa densité est particulièrement forte dans l’hippocampe, le cervelet et le striatum [15]. Le CB2R est présent également dans le système nerveux central [14], mais il est principalement exprimé sur les cellules immunes activées (lymphocytes B et T, macrophages, monocytes, cellules natural killer [NK], etc.) à des concentrations bien plus élevées que le CB1R [14]. La localisation du CB2R expliquerait donc ses fonctions : CB2R module la libération des cytokines et la migration des cellules immunitaires et diminue la présentation d’antigènes, ce qui explique l’intérêt thérapeutique grandissant pour cette cible dans diverses pathologies (cancer, douleur, inflammation) [16]. Ces dernières années, l’existence d’un récepteur CB supplémentaire, le GPR55 (G protein-coupled receptor 55), un récepteur couplé aux protéines G orphelin activé par des cannabinoïdes atypiques ne se liant ni au CB1R ni au CB2R, et par certains EC (AEA, virodhamine), a été suggérée, mais les recherches sur ce récepteur ne sont qu’à leur balbutiement [17]1 (➜). D’autre part, les EC régulent également le récepteur TRPV1 (transient receptor potential channel vanilloid subfamily, member 1), offrant ainsi une opportunité supplémentaire pour le développement de nouvelles thérapies basées sur les EC [18].
(➜) Voir m/s n° 10, vol. 28, octobre 2012
Potentiel thérapeutique des agonistes sélectifs du récepteur CB2
La localisation du CB2R suggère un rôle important de ce récepteur dans le contrôle de l’homéostasie du système immunitaire (production de cytokines pro- et anti-inflammatoires, migration, prolifération et activation de cellules immunitaires) [19]. L’absence d’immunomodulation induite par les cannabinoïdes chez des souris dépourvues du CB2R, souligne le rôle de ce récepteur dans de nombreuses maladies, telles que les maladies inflammatoires, les maladies neurodégénératives (SEP, maladie d’Alzheimer, maladie de Parkinson, troubles neurocognitifs associés au VIH-1 [virus de l’immunodéficience humaine-1]) ou les maladies inflammatoires chroniques de l’intestin (MICI), mais également dans l’athérosclérose [20], la douleur, les cancers, l’ostéoporose ou les maladies du foie par exemple [21]. Dans cette revue, nous n’évoquerons que le potentiel anti-inflammatoire, analgésique et anticancéreux des agonistes sélectifs du CB2R.
Potentiel anti-inflammatoire des agonistes CB2 sélectifs
Les maladies inflammatoires sont induites par une réaction du système immunitaire à une agression. Or, la surexpression du CB2R serait essentielle au contrôle de cette inflammation [22].
Dans les MICI, qui regroupent la maladie de Crohn et la rectocolite hémorragique, une activation irrégulière du système immunitaire est observée, caractérisée par une infiltration prononcée de neutrophiles dans les lésions des muqueuses intestinales, accompagnée de nécroses cellulaires et d’ulcérations de l’épithélium [23]. Le système endocannabinoïde a été identifié dans le tractus gastrointestinal et les CB2R inhiberaient la motilité intestinale et l’inflammation (Figure 1) [24]. Chez les patients souffrant de MICI et dans des modèles animaux de ces maladies, une surexpression des récepteurs CB et une augmentation du taux d’EC ont été observées, résultant d’une réponse adaptative pour rétablir l’homéostasie locale. Des études in vivo et in vitro soulignent le rôle bénéfique du CB2R dans l’inflammation intestinale : des agonistes sélectifs (JWH-015) du récepteur inhibent la production d’IL-8 (interleukine-8, cytokine pro-inflammatoire) dans les cellules humaines HT-29, provenant d’un adénocarcinome colorectal. Cet effet n’est pas observé avec des agonistes sélectifs du CB1R, et il est bloqué par des antagonistes du CB2R [25]. Le système endocannabinoïde, en particulier CB2R, joue également un rôle dans la guérison et la protection intestinales lors d’une inflammation. En effet, il a été montré que JWH-133, un agoniste sélectif du CB2R, diminue l’inflammation du côlon dans un modèle de colite induite par le dextran sulfate de sodium [22].
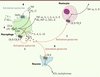 |
Figure 1. Implication des récepteurs CB dans le contrôle de l’inflammation. A. Dans les macrophages, les lipopolysaccharides (LPS), facteurs pathogènes, activent de nombreuses voies de signalisation via le complexe CD14/toll-like receptor 4 (TLR4). Les LPS induisent l’expression de cytokines pro-inflammatoires (TNF-α, IL-1, IL-6, IL-8), de chémokines (MIP-2, CXCL-8), de monoxyde d’azote (NO), de médiateurs lipidiques (prostaglandines [PG], leukotriènes [LT], facteurs d’activation plaquettaire [PAF]), ainsi que la libération d’AEA. B. Les PAF activent de façon autocrine les récepteurs spécifiques (PAFR) des macrophages et stimulent la synthèse de 2-AG. C. L’AEA, médiateur autocrine sur les macrophages et paracrine sur les mastocytes, inhibe la production de cytokines pro-inflammatoires et chémokines via l’activation des CB1R et CB2R, diminuant l’infiltration des leucocytes et l’inflammation. D. L’activation paracrine des CB1R des neurones extrinsèques et intrinsèques inhibe la libération d’acétylcholine (ACh) et de tachykinines respectivement, inhibant ainsi la motilité intestinale [26]. 2-AG : 2-arachidonoylglycérol. |
Malgré sa prédominance périphérique, le CB2R est également surexprimé dans les neurones et les cellules gliales lors d’une inflammation induite par les maladies neurodégénératives chroniques [2]. L’activation du CB2R diminue la production d’agents neurotoxiques immunomodulateurs, incluant les cytokines pro-inflammatoires (NO [oxyde nitrique], TNF-α [tumor necrosis factor-α], IL-1 et IL-6), mais l’activité anti-inflammatoire du CB2R peut également être due à l’activation de cytokines anti-inflammatoires (IL-4, IL-10), qui rétablit ainsi la balance entre cytokines pro- et anti-inflammatoires (Figure 2). Des études dans différents modèles in vitro et in vivo ont également mis en évidence le rôle neuroprotecteur des récepteurs CB, notamment du CB2R.
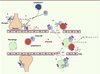 |
Figure 2. Implication du CB2R dans la neuroinflammation. A. Les CB2R sont surexprimés dans les microglies lors de maladies neurodégénératives. Ces cellules gliales s’activent alors pour neutraliser les dommages neuronaux. B. Lors de la progression de la maladie, la barrière hémato-encéphalique (BHE) se fragilise et les macrophages sanguins, les lymphocytes B et les cellules NK expriment à leur tour le CB2R. C. L’activation de ces récepteurs engendre une action anti-inflammatoire en diminuant la libération de cytokines pro-inflammatoires (NO, TNF-α, IL-1, IL-6) et en activant la libération de cytokines anti-inflammatoires (IL-4, IL-10). D. L’activation des CB2R favorise également le recrutement de cellules immunitaires (lymphocytes, cellules NK) de la circulation sanguine suite à la fragilisation de la BHE, initiant ainsi la réponse neuroinflammatoire. E. Finalement, la stimulation des CB2R réprime l’expression du récepteur CD40 induite par l’IFN-γ (interféron) et inhibe également la production microgliale de TNF-α et de NO induite par l’activation de CD40 par l’IFN-γ [27]. |
L’inflammation est l’une des réponses immunitaires les plus fréquentes dans l’organisme et est impliquée dans de nombreuses pathologies. De nombreuses études soutiennent le rôle important du CB2R dans le maintien de l’homéostasie durant une inflammation et font de ce récepteur une cible attractive pour les nouvelles approches thérapeutiques anti-inflammatoires dans le traitement des MICI, des maladies neurodégénératives, mais également l’athérosclérose, l’arthrite rhumatoïde ou encore l’ischémie-reperfusion [28].
Potentiel analgésique des agonistes sélectifs du CB2R
L’utilisation du chanvre dans le traitement de la douleur est connue depuis l’antiquité et, de nos jours, l’efficacité des cannabinoïdes dans le traitement de la douleur aiguë ou chronique a été démontrée. En effet, l’AEA, le THC et les cannabinoïdes synthétiques (CP-55,940, WIN 55,212-2) seraient efficaces en cas de douleur aiguë d’origine chimique (acide acétique, formaldéhyde) ou mécanique (pression) [29]. La coadministration de cannabinoïdes et d’anti-inflammatoires non stéroïdiens provoquerait une synergie d’action en potentialisant l’effet antinociceptif observé, et l’implication des CB2R dans le phénomène d’hyperalgésie a été récemment soulignée [30]. Ainsi, dans un modèle murin d’hypersensibilité viscérale, l’administration d’agonistes du CB2R induit des effets antalgiques protecteurs [31]. Par ailleurs, les agonistes des récepteurs CB libèreraient aussi des opioïdes endogènes et déclencheraient une interaction fonctionnelle entre le SEC et le système opioïde responsable de la modulation des réponses antalgiques (Figure 3) [32]. Ils empêcheraient le développement d’une tolérance aux opiacés ; leur utilisation en tant que complément ou substitut aux opiacés dans le traitement de la douleur chronique est de plus en plus fréquente [32].
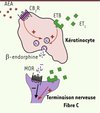 |
Figure 3. Implication du CB2R dans la douleur. En réponse à une lésion cutanée, les kératinocytes sécrètent des neuropeptides (β-endorphine, endothéline-1 [ET1]) qui modulent l’activité des fibres C afférentes. La β-endorphine exerce une action antinociceptive via le récepteur MOR (μ-opioid receptor) des nocicepteurs, alors que l’ET1 exerce une action pronociceptive via le récepteur ETA. L’ET1 active également le récepteur ETB et déclenche ainsi un rétrocontrôle négatif en stimulant la libération de β-endorphine, également favorisée par activation du CB2R des kératinocytes par l’AEA [33]. |
Potentiel anticancéreux des agonistes sélectifs des CB2R
De nombreux travaux révèlent l’implication du système endocannabinoïde dans le cancer. L’activité antinéoplasique du THC a été montrée avant même que le SEC n’ait été découvert. Les EC (AEA, 2-AG) inhibent de façon endogène la prolifération cellulaire cancéreuse. Les récepteurs des cannabinoïdes sont surexprimés de manière significative dans certains cancers, et l’activation in vitro de ces récepteurs réduirait la prolifération cellulaire dans différentes tumeurs (sein, prostate, colorectal, gliome, mélanome, leucémie) [34]. Les recherches sur les cannabinoïdes ont considérablement progressé ces dernières décennies, révélant leur rôle potentiel comme agents anticancéreux. La majorité des travaux se focalisent sur le rôle des cannabinoïdes dans la régulation de la croissance et l’apoptose des cellules tumorales, mais l’impact des cannabinoïdes dans d’autres mécanismes contribuant à l’acquisition d’un phénotype tumoral plus agressif (angiogenèse, migration cellulaire, invasion et métastase) doit être soulignée (Figure 4) [3]. Ainsi, l’activation des CB2R par un agoniste sélectif JWH-133 induirait une régression tumorale (dans le modèle de cellules de gliomes C6 greffées à des souris nude athymiques) [35] ; elle serait responsable de l’action anti-angiogénique de ce composé dans les gliomes malins (via l’inhibition directe de la migration cellulaire endothéliale vasculaire et la diminution de l’expression des facteurs pro-angiogéniques VEGF [vascular endothelial growth factor] et angiopoïétine-2) [35] et favoriserait l’apoptose via la synthèse accrue de novo de céramide [3].
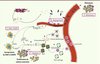 |
Figure 4. Implication des récepteurs CB dans le cancer. A. Les leucocytes de la circulation sanguine migrent vers la tumeur suite à la production de chémokines, formant ainsi des macrophages associés aux tumeurs (MAT). B. Les MAT sécrètent de nombreuses cytokines inflammatoires (IL-1β, IL-6, TNF-α), qui stimulent la prolifération des cellules tumorales, la surexpression du gène codant pour le VEGF, l’induction de COX-2 et de PGE2. C. VEGF et PGE2 agissent alors sur les cellules endothéliales causant l’angiogenèse. D. Les MAT favorisent également l’échappement tumoral de la surveillance immunitaire en produisant des immunotoxines qui inactivent les cellules dendritiques. E. Les fibroblastes associés aux carcinomes (FAC) sécrètent des taux élevés de SDF-1 (ou CXCL12), chémokine favorisant la progression de la tumeur via le recrutement de cellules endothéliales progénitrices (CEP) circulantes, la stimulation de l’angiogenèse, la stimulation directe des cellules tumorales et la migation des cellules métastatiques dans les organes spécifiques. Les CB diminuent les taux de VEGF et VEGFR surexprimés dans les tumeurs, les métastases et les cellules endothéliales vasculaires, réduisant ainsi l’angiogenèse, la migration des cellules cancéreuses et les métastases [3]. |
 |
Figure 5. Structures chimiques du THC et de quelques agonistes du CB2R. |
Conclusion
Ces dernières décennies, l’implication du SEC dans de nombreuses fonctions physiologiques a été mise en évidence. Ce système maintient l’homéostasie dans des conditions physiologiques normales et pathologiques et joue un rôle protecteur dans de nombreuses maladies. En effet, dans certains cancers ou maladies inflammatoires, la concentration et l’expression accrues des EC diminueraient les symptômes et la progression de la pathologie. Ainsi, la modulation de ce système (modification des taux d’EC et activation des récepteurs CB) semble être une stratégie thérapeutique intéressante dans de nombreuses pathologies (inflammation, cancer, douleur). Dans ce cadre, le développement de cannabinoïdes synthétiques apparaît très prometteur [36]. Ces composés présentent de nombreuses vertus, mais étant donné leur développement récent, peu de données concernant les effets secondaires existent actuellement. Néanmoins, les effets néfastes du cannabis semblent être moins graves que ceux de l’alcool en termes d’effets neuropsychologiques et somatiques, d’accidents et de violence. Il est cependant admis que les effets centraux indésirables (effets psychiatriques, cognitifs et dépendance) du cannabis résulteraient de l’activation du CB1R ; par conséquent, le développement d’agonistes sélectifs du CB2R semble fondamental.
Liens d’intérêt
Les auteurs déclarent n’avoir aucun lien d’intérêt concernant les données publiées dans cet article.
Voir le numéro thématique que médecine/sciences a consacré aux récepteurs couplés aux protéines G en octobre 2012.
Références
- Gaoni Y, Mechoulam R. Isolation, structure and partial synthesis of an active constituent of hashish. J Am Chem Soc 1964 ; 86 : 1646–1647. [CrossRef] [Google Scholar]
- Scotter EL, Abood ME, Glass M. The endocannabinoid system as a target for the treatment of neurodegenerative disease. Br J Pharm 2010 ; 160 : 480–498. [CrossRef] [Google Scholar]
- Fogli S, Breschi MC. The molecular bases of cannabinoid action in cancer. Cancer Therapy 2008 ; 6 : 103–116. [Google Scholar]
- MacCoun R, Reuter P. Evaluating alternative cannabis regimes. Br J Psychiatry 2001 ; 178 : 123–128. [CrossRef] [PubMed] [Google Scholar]
- Hazekamp A, Grotenhermen F. Review on clinical studies with cannabis and cannabinoids 2005–2009. cannabinoids 2010 ; 5 : 1–21. [Google Scholar]
- Piomelli D. The molecular logic of endocannabinoid signalling. Nat Rev Neurosci 2003 ; 4 : 873–884. [CrossRef] [PubMed] [Google Scholar]
- Giuffrida R, Beltramo M, Piomelli D. Mechanism of endocannabinoid inactivation: biochemistry and pharmacology. J Pharm Exp Ther 2001 ; 298 : 7–14. [Google Scholar]
- Steffens S, Mach M. Vers un usage thérapeutique de ligands sélectifs du récepteur cannabinoïde 2 dans l’athérosclérose. Med Sci (Paris) 2006 ; 1 : 7–9. [CrossRef] [EDP Sciences] [Google Scholar]
- Vandevoorde S, Lambert DM. Focus on the three key enzymes hydrolysing endocannabinoids as new drug targets. Curr Pharm Des 2005 ; 18 : 2647–2668. [CrossRef] [Google Scholar]
- Di Marzo V. Targeting the endocannabinoid system: to enhance or reduce? Nat Rev 2008 ; 7 : 438–455. [CrossRef] [Google Scholar]
- Munro LA, Lolait SJ, Brownstein MJ, et al. Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature 1990 ; 346 : 561–564. [CrossRef] [PubMed] [Google Scholar]
- Munro S, Thomas KL, Abu-Shaar M. Molecular characterization of a peripheral receptor for cannabinoids. Nature 1993 ; 365 : 61–65. [CrossRef] [PubMed] [Google Scholar]
- Onaivi ES. An endocannabinoid hypothesis of drug reward and drug addiction. Ann N Y Acad Sci 2008 ; 1139 : 412–421. [CrossRef] [PubMed] [Google Scholar]
- Galiègue S, Mary S, Marchand J, et al. Expression of central and peripheral cannabinoid receptors in human immune tissues and leukocyte subpopulations. Eur J Biochem 1995 ; 232 : 54–61. [CrossRef] [PubMed] [Google Scholar]
- Van Sickle MD, Duncan M, Kingsley PJ, et al. Identification and functional characterization of brainstem cannabinoid CB2 receptors. Science 2005 ; 310 : 329–332. [CrossRef] [PubMed] [Google Scholar]
- Yang P, Wang L, Xie X-Q. Latest advances in novel cannabinoid CB2 ligands for drug abuse and their therapeutic potential. Future Med Chem 2012 ; 4 : 187–204. [CrossRef] [PubMed] [Google Scholar]
- Sharir H, Console-Bram L, Mundy C, et al. The endocannabinoids anandamide and virodhamine modulate the activity of the candidate cannabinoid receptor GPR55. J Neuroimmune Pharmacol 2012 ; 7 : 56–65. [CrossRef] [Google Scholar]
- Di Marzo V, De Petrocellis L. Endocannabinoids as regulators of transient receptor potential (TRP) channels: A further opportunity to develop new endocannabinoid-based therapeutic drugs. Curr Med Chem 2010 ; 17 : 1430–49. [CrossRef] [PubMed] [Google Scholar]
- Klegeris A, Bissonnette CJ, McGeer PL. Reduction of human monocytic cell neurotoxicity and cytokine secretion by ligands of the cannabinoid-type CB2 receptor. Br J Pharmacol 2003 ; 139 : 775–786. [CrossRef] [PubMed] [Google Scholar]
- Venance L, Maldonado R. Manzoni. Le système endocannabinoïde central. Med Sci Paris 2004 ; 1 : 45–53. [Google Scholar]
- Pacher P, Mechoulam R. Is lipid signaling through cannabinoid 2 receptors part of a protective system? Prog Lipid Res 2011 ; 50 : 193–211. [CrossRef] [PubMed] [Google Scholar]
- Wright K, Rooney N, Feeney M, et al. Differential expression of cannabinoid receptors in the human colon: cannabinoids promote epithelial wound healing. Gastroenterology 2005 ; 129 : 437–453. [PubMed] [Google Scholar]
- Podolsky DK. Inflammatory bowel disease. N Engl J Med 2002 ; 347 : 417–429. [CrossRef] [PubMed] [Google Scholar]
- Di Marzo V, Izzo AA. Endocannabinoid overactivity and intestinal inflammation. Gut 2006 ; 55 : 1373–1376. [CrossRef] [PubMed] [Google Scholar]
- Ihenetu K, Molleman A, Parsons ME, et al. Inhibition of interleukin-8 release in the human colonic epithelial cell line HT-29 by cannabinoids. Eur J Pharmacol 2003 ; 458 : 207–215. [CrossRef] [PubMed] [Google Scholar]
- Kunos G, Pacher P. Cannabinoids cool the intestine. Nat Med 2004 ; 10 : 678–679. [CrossRef] [PubMed] [Google Scholar]
- Ehrhart J, Obregon D, Mori T, et al. Stimulation of cannabinoid receptor 2 (CB2) suppresses microglial activation. J Neuroinflam 2005 ; 2 : 29. [CrossRef] [Google Scholar]
- Leleu-Chavain N, Body-Malapel M, Spencer J, et al. Recent advances in the development of selective CB2 agonists as promising anti-inflammatory agents. Curr Med Chem 2012 ; 19 : 3457–3474. [CrossRef] [PubMed] [Google Scholar]
- Calignano A, La Rana G, Giuffrida A, et al. Control of pain initiation by endogenous cannabinoids. Nature 1998 ; 394 : 277–281. [CrossRef] [PubMed] [Google Scholar]
- Guindon J, Beaulieu P. Antihyperlagesic effects of local injections of anandamide, ibuprofenn rofecoxib and their combinations in a model of neuropathic pain. Neuropharmacology 2006 ; 50 : 814–823. [CrossRef] [PubMed] [Google Scholar]
- Kikuchi A, Ohashi K, Sugie Y, et al. Pharmacological evaluation of a novel cannabinoid 2 (CB2) ligand, PF-03550096, in vitro and in vivo by using a rat model of visceral hypersensitivity. J Pharmacol Sci 2008 ; 106 : 219–224. [CrossRef] [PubMed] [Google Scholar]
- Lucas P. Cannabis as an adjunct to or substitute for opiates in the treatment of chronic pain. J Psychoactive Drugs 2012 ; 44 : 125–133. [CrossRef] [PubMed] [Google Scholar]
- Calvino B. Les kératinocytes : des cellules non neuronales impliquées dans la douleur. Douleurs Évaluation - Diagnostic - Traitement 2010 ; 11 : 120–130. [CrossRef] [Google Scholar]
- Sarfaraz S, Afaq F, Adhami VM, et al. Cannabinoid receptor as a novel target for the treatment of prostate cancer. Cancer Res 2005 ; 65 : 1635–1641. [CrossRef] [PubMed] [Google Scholar]
- Blazquez C, Casanova ML, Planas A, et al. Inhibition of tumor angiogenesis by cannabinoids. FASEB J 2003 ; 17 : 529–531. [PubMed] [Google Scholar]
- Fatima Teixeira-Clerc, Boris Julien, Pascale Grenard, et al. Les antagonistes du récepteur CB1 des cannabinoïdes : une nouvelle approche pour le traitement de la fibrose hépatique. Med Sci (Paris) 2006 ; 22 : 683–685. [CrossRef] [EDP Sciences] [PubMed] [Google Scholar]
Liste des figures
|
Figure 1. Implication des récepteurs CB dans le contrôle de l’inflammation. A. Dans les macrophages, les lipopolysaccharides (LPS), facteurs pathogènes, activent de nombreuses voies de signalisation via le complexe CD14/toll-like receptor 4 (TLR4). Les LPS induisent l’expression de cytokines pro-inflammatoires (TNF-α, IL-1, IL-6, IL-8), de chémokines (MIP-2, CXCL-8), de monoxyde d’azote (NO), de médiateurs lipidiques (prostaglandines [PG], leukotriènes [LT], facteurs d’activation plaquettaire [PAF]), ainsi que la libération d’AEA. B. Les PAF activent de façon autocrine les récepteurs spécifiques (PAFR) des macrophages et stimulent la synthèse de 2-AG. C. L’AEA, médiateur autocrine sur les macrophages et paracrine sur les mastocytes, inhibe la production de cytokines pro-inflammatoires et chémokines via l’activation des CB1R et CB2R, diminuant l’infiltration des leucocytes et l’inflammation. D. L’activation paracrine des CB1R des neurones extrinsèques et intrinsèques inhibe la libération d’acétylcholine (ACh) et de tachykinines respectivement, inhibant ainsi la motilité intestinale [26]. 2-AG : 2-arachidonoylglycérol. |
| Dans le texte | |
|
Figure 2. Implication du CB2R dans la neuroinflammation. A. Les CB2R sont surexprimés dans les microglies lors de maladies neurodégénératives. Ces cellules gliales s’activent alors pour neutraliser les dommages neuronaux. B. Lors de la progression de la maladie, la barrière hémato-encéphalique (BHE) se fragilise et les macrophages sanguins, les lymphocytes B et les cellules NK expriment à leur tour le CB2R. C. L’activation de ces récepteurs engendre une action anti-inflammatoire en diminuant la libération de cytokines pro-inflammatoires (NO, TNF-α, IL-1, IL-6) et en activant la libération de cytokines anti-inflammatoires (IL-4, IL-10). D. L’activation des CB2R favorise également le recrutement de cellules immunitaires (lymphocytes, cellules NK) de la circulation sanguine suite à la fragilisation de la BHE, initiant ainsi la réponse neuroinflammatoire. E. Finalement, la stimulation des CB2R réprime l’expression du récepteur CD40 induite par l’IFN-γ (interféron) et inhibe également la production microgliale de TNF-α et de NO induite par l’activation de CD40 par l’IFN-γ [27]. |
| Dans le texte | |
|
Figure 3. Implication du CB2R dans la douleur. En réponse à une lésion cutanée, les kératinocytes sécrètent des neuropeptides (β-endorphine, endothéline-1 [ET1]) qui modulent l’activité des fibres C afférentes. La β-endorphine exerce une action antinociceptive via le récepteur MOR (μ-opioid receptor) des nocicepteurs, alors que l’ET1 exerce une action pronociceptive via le récepteur ETA. L’ET1 active également le récepteur ETB et déclenche ainsi un rétrocontrôle négatif en stimulant la libération de β-endorphine, également favorisée par activation du CB2R des kératinocytes par l’AEA [33]. |
| Dans le texte | |
|
Figure 4. Implication des récepteurs CB dans le cancer. A. Les leucocytes de la circulation sanguine migrent vers la tumeur suite à la production de chémokines, formant ainsi des macrophages associés aux tumeurs (MAT). B. Les MAT sécrètent de nombreuses cytokines inflammatoires (IL-1β, IL-6, TNF-α), qui stimulent la prolifération des cellules tumorales, la surexpression du gène codant pour le VEGF, l’induction de COX-2 et de PGE2. C. VEGF et PGE2 agissent alors sur les cellules endothéliales causant l’angiogenèse. D. Les MAT favorisent également l’échappement tumoral de la surveillance immunitaire en produisant des immunotoxines qui inactivent les cellules dendritiques. E. Les fibroblastes associés aux carcinomes (FAC) sécrètent des taux élevés de SDF-1 (ou CXCL12), chémokine favorisant la progression de la tumeur via le recrutement de cellules endothéliales progénitrices (CEP) circulantes, la stimulation de l’angiogenèse, la stimulation directe des cellules tumorales et la migation des cellules métastatiques dans les organes spécifiques. Les CB diminuent les taux de VEGF et VEGFR surexprimés dans les tumeurs, les métastases et les cellules endothéliales vasculaires, réduisant ainsi l’angiogenèse, la migration des cellules cancéreuses et les métastases [3]. |
| Dans le texte | |
|
Figure 5. Structures chimiques du THC et de quelques agonistes du CB2R. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.