")
")
| Issue |
Med Sci (Paris)
Volume 28, Number 4, Avril 2012
|
|
|---|---|---|
| Page(s) | 409 - 415 | |
| Section | M/S Revues | |
| DOI | https://doi.org/10.1051/medsci/2012284019 | |
| Published online | 25 avril 2012 | |
Le rôle des kinases Mek1 et Mek2 dans la formation de la barrière hématoplacentaire chez la souris
Mek1 and Mek2 functions in the formation of the blood placental barrier
Centre de recherche en cancérologie de l’université Laval, centre de recherche du centre hospitalier universitaire de Québec, l’Hôtel-Dieu de Québec, 9, rue McMahon, Québec G1R 2J6, Canada
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
**
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
***
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Résumé
La voie de signalisation cellulaire ERK/MAPK (extracellular signal-regulated kinase/mitogen-activated protein kinase) est impliquée dans plusieurs fonctions cellulaires. Chez la souris, l’invalidation de gènes codant pour des membres de cette voie est souvent associée à une mort embryonnaire consécutive à un développement anormal du placenta. Ce dernier est essentiel aux échanges nutritionnels et gazeux entre les circulations maternelle et embryonnaire, ainsi qu’à l’évacuation des déchets métaboliques. Ces échanges s’effectuent sans contact direct entre les deux circulations. Chez la souris, la barrière hématoplacentaire est constituée d’une triple couche de cellules trophoblastiques et de cellules endothéliales de l’embryon. MEK1 (ou MAP/ERK kinase-1) et MEK2 sont des kinases à double spécificité, phosphorylant les résidus sérine/thréonine et tyrosine et responsables de l’activation des kinases ERK1 et ERK2. L’inactivation génique de Mek1 produit une malformation placentaire résultant de défauts de prolifération et de différenciation des cellules trophoblastiques et entraînant un retard sévère du développement du placenta et la mort de l’embryon. Bien que les mutants Mek2-/- survivent sans aucun phénotype apparent, les mutants doubles hétérozygotes Mek1+/-Mek2+/- meurent au cours de la gestation de malformation placentaire. L’ensemble de ces données souligne le rôle crucial de la voie ERK/MAPK dans la formation des structures extraembryonnaires.
Abstract
The ERK/MAPK signaling pathway is involved in several cellular functions. Inactivation in mice of genes encoding members of this pathway is often associated with embryonic death resulting from abnormal placental development. The placenta is essential for nutritional and gaseous exchanges between maternal and embryonic circulations, as well as for the removal of metabolic wastes. These exchanges take place without direct contact between the two circulations. In mice, the hematoplacental barrier consists in a triple layer of trophoblast cells and endothelial cells of the embryo. MEK1 and MEK2 are double specificity serine-threonine/tyrosine kinases responsible for the activation of ERK1 and ERK2. Mek1 inactivation results in placental anomalies due to trophoblast cell proliferation and differentiation defects leading to severe delays in the development of placenta and causing the death of the embryo. Although Mek2-/- mutant mice survived without any apparent phenotype, double heterozygous Mek1+/-Mek2+/- mutants die during gestation from placental malformations. Together, these data emphasize the crucial role of the ERK/MAPK cascade in the formation of extraembryonic structures.
© 2012 médecine/sciences – Inserm / SRMS
Fonctions essentielles de la voie ERK/MAPK lors du développement placentaire murin
La voie de signalisation ERK/MAPK
Les voies de signalisation MAP kinase (MAPK) sont composées d’une succession de protéines kinases pouvant relayer des signaux extracellulaires aux différentes cibles intracellulaires localisées dans la membrane plasmique, le cytoplasme, le cytosquelette ou le noyau [1, 2]. Plusieurs cascades de signalisation MAPK ont été décrites chez les vertébrés. La voie classique ERK/MAPK constitue la voie majeure dans la réponse aux facteurs de croissance. Cette voie utilise les kinases régulées par les signaux extracellulaires, ERK1 et ERK2, ainsi que les kinases MEK1 et MEK2 à double spécificité (sérine/thréonine et tyrosine), qui activent ERK1 et ERK2 en réponse à la liaison de facteurs de croissance à leur récepteur. La voie ERK/MAPK est impliquée dans la détermination cellulaire chez plusieurs espèces. Chez les mammifères, cette cascade est impliquée dans la prolifération, la différenciation, la migration, la survie et la mort cellulaires [3, 4].
La voie d’activation ERK/MAPK débute par la liaison de facteurs de croissance aux récepteurs membranaires à activité tyrosine kinase, tels les récepteurs du FGF (fibroblast growth factor), de l’EGF (epidermal growth factor) et du PDGF (platelet derived growth factor). Ces récepteurs s’activent par une autophosphorylation suivie du recrutement d’une protéine adaptatrice comme GRB2 (growth factor receptor-bound protein 2) et l’interaction avec SOS-1 (son of sevenless homolog 1), un facteur d’échange de GDP/GTP de la petite protéine G RAS. SOS-1 permet le relargage du GDP associé à RAS qui peut alors fixer le GTP et s’activer. RAS activé permet le recrutement et l’activation de la kinase proto-oncogène RAF. Par la suite, les protéines kinases RAF, MEK et ERK s’activent séquentiellement par phosphorylation [5]. À ce jour, les substrats connus des ERK sont nombreux et incluent des protéines du cytosol, du cytosquelette, de la membrane plasmique ainsi que des facteurs de transcription. L’abondance de substrats reflète d’ailleurs l’importance de la voie dans un grand nombre de processus cellulaires [2, 6]. Le rôle de la voie ERK/MAPK lors du développement placentaire a été largement démontré chez la souris par la caractérisation de mutants pour des gènes associés à l’activation de la voie. Les mutants Erk2, Fgfr2, Gab1 (GRB2-associated-binding protein 1), Mek1, Sos1, b-Raf, Raf-1 et le mutant hypomorphe pour Grb2 décèdent entre les jours 10,5 et 11,5 de la gestation et présentent tous des anomalies de développement placentaire (Tableau I) [7–19]. Pour l’ensemble de ces mutants, aucune anormalité morphologique n’est observable chez l’embryon, mais un retard de croissance est noté [7, 20].
Phénotype placentaire des souris mutantes pour des protéines impliquées dans la voie ERK/MAPK.
Le développement du placenta chez la souris
Chez la souris, le développement du placenta s’effectue en deux phases. Vers le 8,5e jour de gestation, il y a liaison du mésoderme allantoïde - qui est à l’origine de l’élongation de la ligne primitive - à l’épithélium chorionique. Par la suite et en réponse à cette interaction, une série de ramifications entre ces deux tissus va façonner l’aire de surface nécessaire aux échanges, qui constitue la région du labyrinthe [21]. À terme, le placenta est formé de trophoblastes dérivant de l’ectoderme extraembryonnaire et du cône ectoplacentaire, d’une vascularisation fœtale provenant du mésoderme allantoïde dérivé de l’embryon, et du sang maternel qui s’épanche de la vascularisation maternelle dans les sinus maternels du labyrinthe. L’intégrité du tissu placentaire peut donc être affectée par des défauts touchant les tissus embryonnaires, les tissus maternels ou les cellules trophoblastiques du placenta. De plus, des études génétiques ont démontré l’importance des interactions entre ces différents constituants afin d’établir un placenta fonctionnel.
Les cellules trophoblastiques ont un rôle primordial de soutien et elles sont activement impliquées dans la vascularisation du placenta [7]. Trois populations trophoblastiques distinctes sont présentes dans le placenta mature. La partie la plus interne est constituée des trophoblastes du labyrinthe (Figure 1A). Elle est juxtaposée à une couche intermédiaire formée de spongiotrophoblastes. La partie la plus externe comprend les cellules trophoblastiques géantes en contact avec les tissus maternels. Les spongiotrophoblastes ont un rôle structural et sécrètent des facteurs antiangiogéniques qui empêcheraient la croissance des vaisseaux maternels dans la partie fœtale du placenta [22]. Par ailleurs, les cellules trophoblastiques géantes exercent des fonctions précoces qui favorisent l’implantation et l’invasion de l’utérus par l’embryon. Par la suite, les trophoblastes géants, via leurs fonctions endocrines, promeuvent les adaptations physiologiques de la mère [23]. Les trophoblastes de la couche interne du labyrinthe comprennent les trophoblastes mononucléés sinusoïdaux géants (S-TGC) et une double couche de syncytiotrophoblastes (SynT). Les S-TGC font face aux sinus maternels et sont juxtaposés à la première couche de SynT (SynT-I), ellemême accolée à une deuxième couche de SynT (SynT-II) adjacente à l’endothélium fœtal. Les cellules sanguines maternelles et les érythrocytes fœtaux sont donc séparés par une triple couche de cellules trophoblastiques du labyrinthe et par les cellules endothéliales fœtales (Figure 1B) [20, 24]. Les S-TGC sont un sous-type de trophoblastes géants pourvus de fonctions endocrines. Les SynT-I et SynT-II sont des cellules postmitotiques multinucléées formées par fusion cellulaire et possédant des fonctions de transport [20, 25]. Ainsi, des anomalies affectant les cellules trophoblastiques ou les cellules endothéliales issues de l’allantoïde peuvent mener à des problèmes de vascularisation placentaire.
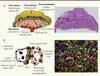 |
Figure 1. Structure du placenta de la souris : organisation des différentes couches cellulaires de la région du labyrinthe de souris. A. Représentation schématique du placenta de souris (panneau de gauche) et coupe histologique d’un placenta d’un embryon au 12,5e jour de gestation (panneau de droite). La ligne pointillée sépare les parties maternelle et embryonnaire. B. Schéma de l’organisation de la triple couche de trophoblastes et de l’endothélium fœtal dans la région du labyrinthe permettant la formation de la barrière hématoplacentaire (panneau de gauche). Visualisation de la double couche de syncytiotrophoblastes par immunofluorescence avec des anticorps contre les transporteurs de monocarboxylate MCT1 (en rouge) et MCT4 (en vert) (panneau de droite) [24]. MCT1 et MCT4 sont utilisés comme marqueurs spécifiques de la membrane apicale des SynT-I et de la membrane basale des SynT-II, respectivement. Dans cette expérience, les noyaux sont marqués en bleu par une coloration au DAPI. Les érythrocytes maternels sont détectés en orange (du fait de l’autofluorescence), alors que les érythrocytes fœtaux ont un noyau bleu et un cytoplasme verdâtre. Les érythrocytes et les sinus maternels sont délimités par la couche de SynT-I (en rouge), alors que les érythrocytes et les cellules endothéliales des vaisseaux sanguins fœtaux sont inclus à l’intérieur d’une couche de SynT-II (en vert). Dans le vaisseau sanguin fœtal indiqué (VSF), une cellule endothéliale (CE) accolée sur la partie interne de la couche de SynT-II définie par le marquage MCT4 est visible. Les circulations sanguines fœtale et maternelle sont séparées par trois couches étanches constituées des cellules endothéliales vasculaires fœtales, des SynT-I et des SynT-II. CE, cellule endothéliale ; EF, érythrocyte fœtal ; EM, érythrocyte maternel ; SM, sinus maternel ; SynT-I, syncytiotrophoblaste de type I ; SynT-II, syncytiotrophoblaste de type II ; VSF, vaisseau sanguin fœtal. |
Défauts des lignées trophoblastiques chez les mutants de la voie ERK
Mutations de gènes de la voie de signalisation ERK/MAPK et développement labyrinthique
Les techniques d’agrégation d’embryons tétraploïdes1 et de délétion conditionnelle de gènes chez la souris ont permis de démontrer que la mutation de gènes associés à la voie de signalisation ERK/MAPK (Gab1, Sos1, b-Raf, Mek1, Erk2) entraîne la mort embryonnaire en raison des malformations placentaires résultant de défauts dans les tissus extraembryonnaires [7, 9, 13, 17, 26]. Chez ces différents mutants, le sous-développement de la région du labyrinthe est associé à une incapacité du mésoderme allantoïde à envahir cette région. Les vaisseaux embryonnaires placentaires chez les mutants ne se retrouvent pas à proximité des sinus maternels, ce qui diminue les échanges gazeux et nutritionnels et cause la mort de l’embryon. À titre d’exemple, la région du labyrinthe des placentas Mek1-/- est atrophiée en raison d’une prolifération diminuée et d’une apoptose accrue des trophoblastes. De plus, le labyrinthe est hypovascularisé [12–14]. L’utilisation du facteur de transcription codé par le gène Gcm1 (glial cells missing homolog 1) comme marqueur des SynT-II a montré que ce type cellulaire est correctement spécifié dans les placentas Mek1-/-, mais que les SynT-II demeurent bloqués à la jonction chorio-allantoïde [13]. Il a été proposé que les SynT-II seraient les cellules responsables de l’émergence de la morphogenèse de la région du labyrinthe, ouvrant ainsi le chemin à la migration des cellules endothéliales [21]. L’incapacité des SynT-II à migrer dans la région du labyrinthe expliquerait l’hypovascularisation du placenta des souris Mek1-/-.
Distribution spatiale des protéines MEK1 et MEK2
La distribution spatiale des protéines MEK1 et MEK2 diffère dans le placenta. Alors que la protéine MEK2 est exprimée de façon ubiquitaire, MEK1 est plus fortement exprimée dans les tissus situés entre les sinus maternels et les vaisseaux embryonnaires, possiblement dans les couches trophoblastiques composant la barrière hématoplacentaire [13, 15]. L’analyse des formes phosphorylées de ERK1, ERK2, MEK1 et MEK2 révèle que la voie ERK/MAPK est principalement activée près des sinus maternels (Figure 2). Le profil d’expression des membres de la cascade ERK/MAPK, combiné à des études génétiques, a permis de montrer le rôle prépondérant de cette voie dans les tissus du placenta ayant une origine extraembryonnaire (plutôt que dans les cellules endothéliales) pour l’établissement de la barrière hématoplacentaire. En effet, la délétion spécifique du gène b-Raf dans les cellules endothéliales à l’aide de la lignée transgénique Tie2-Cre ou encore la délétion du gène Mek1 par la recombinase Sox2-Cre dans tous les tissus embryonnaires (incluant l’allantoïde), ne génère aucun défaut placentaire et aucune mort de l’embryon puisque ces gènes demeurent fonctionnels dans les cellules trophoblastiques [13, 17]. Bien qu’il ait été rapporté que l’activation de ERK dans le placenta se produisait près des vaisseaux embryonnaires [17], la majorité des données obtenues à ce jour supportent plutôt un rôle crucial de la voie ERK/MAPK dans les tissus extraembryonnaires pour la formation du placenta.
 |
Figure 2. Activation différentielle des voies ERK et p38 MAPK dans les cellules du labyrinthe. Le réseau vasculaire embryonnaire est révélé à l’aide d’un anticorps anti-CD31 (cluster of differentiation 31) alors que l’activation des voies de signalisation ERK/MAPK et p38/MAPK est mise en évidence avec des anticorps contre les formes phosphorylées de ERK (p-ERK) et de p38 (p-p38), respectivement. Les images sont prises en contraste interférentiel. Les érythrocytes embryonnaires et maternels sont détectables dans les vaisseaux fœtaux (VF) et les sinus maternels (SM), respectivement. Les érythrocytes embryonnaires sont plus gros et nucléés. L’activation de la voie p38/MAPK est restreinte aux vaisseaux sanguins fœtaux alors que l’activation de ERK est localisée en périphérie des sinus maternels. Les flèches noires indiquent les cellules endothéliales vasculaires fœtales et les flèches blanches les trophoblastes bordant les sinus maternels. |
Voie ERK/MAPK et expression du VEGF
La voie ERK/MAPK joue un rôle important dans le maintien de l’expression du VEGF (vascular endothelial growth factor). Chez les souris mutantes b-Raf, l’expression du VEGF est diminuée. En revanche, la délétion spécifique de b-Raf dans les cellules endothéliales n’a pas d’effet sur la formation du placenta, suggérant que le phénotype placentaire observé chez la souris b-Raf-/- est causé par une perte d’expression de VEGF dans les tissus placentaires autres que les cellules endothéliales [17]. Chez les mutants Mek1, l’expression du gène Vegf est fortement augmentée indiquant une hypoxie au niveau du placenta causée par l’absence de développement du labyrinthe [13]. L’augmentation d’expression de Vegf corrèle avec une augmentation de l’activation de p38/MAPK [13]. La voie p38/MAPK est impliquée dans la réponse au VEGF lors de l’angiogenèse et elle est fortement activée dans les cellules endothéliales du placenta en formation (Figure 2) [27]. Ces résultats montrent que l’expression du VEGF est dépendante de b-Raf, mais indépendante de l’activation de ERK par MEK1. Une explication possible serait que MEK2 pourrait compenser la perte de MEK1. Une autre hypothèse serait qu’en absence de MEK1, l’activation de la voie ERK/MAPK serait soutenue puisqu’elle utiliserait uniquement MEK2. En effet, il a été montré que MEK1 est essentielle pour la boucle de rétro-inhibition de la voie par la phosphorylation de l’hétérodimère MEK1-MEK2 par ERK. En absence de MEK1, l’activation de ERK par MEK2 pourrait être moindre, bien que stabilisée [14]. Ainsi, le maintien de l’activation de la voie ERK/MAPK pourrait mener à une augmentation de l’expression de VEGF. Cependant, l’analyse de l’activation de ERK dans les placentas Mek1-/- n’a pour l’instant pas permis de confirmer cette dernière hypothèse [13, 14].
Rôle de la voie ERK/MAPK dans les SynT-II
Les embryons Mek1-/- meurent à mi-gestation alors que les souris Mek2-/- survivent sans aucun phénotype apparent [13, 28]. En revanche, une majorité des mutants Mek1+/-Mek2+/- meurent au stade E14,5 en raison d’anomalies au niveau des trophoblastes, révélant ainsi que Mek2 joue également un rôle lors du développement du placenta. L’haplo-insuffisance de Mek2 lorsqu’un allèle de Mek1 est absent, engendre un sous-développement de la région du labyrinthe et une diminution de la vascularisation. Ce phénotype, semblable à celui des mutants Mek1-/-, est néanmoins moins prononcé et moins pénétrant. De plus, les placentas Mek1+/-Mek2+/- présentent une accumulation de structures anormales composées de cellules multinucléées trophoblastiques géantes (MTG) localisées dans le labyrinthe. Des études génétiques de lignage cellulaire in vivo, utilisant la lignée de souris transgénique Gcm1Cre et le gène rapporteur Rosa26-LacZ, ont permis de montrer que les cellules MTG dérivent des SynT-II [15]. Par ailleurs, l’utilisation de marqueurs cellulaires a révélé que les cellules MTG n’expriment plus le gène Gcm1. Ces résultats suggèrent que la voie ERK/MAPK est impliquée dans le maintien de l’expression de Gcm1, mais non dans l’initiation de sa transcription, comme précédemment démontré chez l’oursin de mer pour le gène spgcm, l’homologue de Gcm1 [29]. En accord avec cette hypothèse, l’activation de la voie ERK/MAPK est perdue dans les cellules MTG alors qu’elle est maintenue dans les SynT des régions exemptes de MTG.
Dans les placentas d’embryons Mek2+/-, la délétion conditionnelle d’un allèle de Mek1 spécifiquement dans les SynT-II est suffisante pour générer des cellules MTG, démontrant l’importance de MEK1 dans les SynT-II lorsque les niveaux d’expression de Mek2 sont diminués [15]. En revanche, le phénotype est moins prononcé que celui observé dans les placentas Mek1+/-Mek2+/-, ce qui suggère un rôle plus large de MEK1 dans d’autres types cellulaires du placenta. Des structures semblables aux cellules MTG ont aussi été rapportées pour les mutants Sos1 et b-Raf [17, 30]. Puisque SOS1 et b-RAF se situent en amont de ERK dans la voie, ces données suggèrent qu’ils pourraient participer à l’activation spécifique de ERK dans les SynT-II afin de promouvoir une différenciation et/ou une polarisation adéquate de ces cellules.
Pparγ et Lbp-1a : cibles potentielles de la voie ERK/MAPK dans la formation du placenta
Les souris mutantes pour les gènes Pparγ (peroxisome proliferator-activated γ) et Lbp-1a (ou upstream binding protein 1) présentent aussi des phénotypes placentaires semblables à ceux qui sont observés chez les mutants Mek [31–33]. PPARγ est un récepteur nucléaire impliqué dans plusieurs fonctions dont la différenciation, la prolifération, le métabolisme tissulaire et l’immunité de l’hôte [34, 35]. L’invalidation de ce gène provoque une mortalité embryonnaire à mi-gestation due à un défaut dans les lignées trophoblastiques [33, 36]. Le labyrinthe des placentas Pparγ-/- est atrophié et sa vascularisation est désorganisée. Les trophoblastes du labyrinthe des placentas d’embryons Pparγ-/- présentent des défauts de différenciation affectant la formation du syncytium. De plus, des structures semblables aux cellules MTG sont présentes dans les placentas des spécimens mutants [36]. Le gène Pparγ est exprimé de façon ubiquitaire au niveau des trophoblastes du labyrinthe. L’étude des placentas Pparγ-/- en microscopie électronique a révélé un déficit dans l’accumulation de gouttelettes lipidiques dans les SynT-II indiquant un problème de différenciation en plus d’une adhésion inadéquate à l’endothélium [33]. L’analyse des cellules souches trophoblastiques Pparγ-/- a aussi mis en évidence le rôle de PPARγ dans le contrôle de la transcription des gènes Muc1 codant pour la glycoprotéine mucine 1 et Gcm1 codant pour le chorion-specific transcription factor 1 [31]. Gcm1 est exprimé spécifiquement dans les SynT-II et est impliqué dans leur différenciation [37]. Pour sa part, l’expression de Muc1 est restreinte à la bordure des sinus maternels (SynT-I ou S-TGC), ce qui suggère un rôle pour PPARγ dans les SynT-I, SynT-II et/ou S-TGC. Par ailleurs, le phénotype placentaire partiel des mutants Muc1-/- suggère que les fonctions essentielles de PPARγ sont relayées par d’autres cibles lors de la formation du placenta [38].
PPARγ est une cible connue de la voie ERK/MAPK [34, 39, 40]. La phosphorylation de PPARγ par ERK contribue à la réduction de son activité transcriptionnelle induite par les facteurs de croissance [39]. De plus, il a été démontré que MEK1 peut interagir directement avec PPARγ et induire son transport dans le cytoplasme indépendamment de la phosphorylation de PPARγ par ERK [34]. Ce transport nucléaire, réalisé par l’intermédiaire de MEK1, atténue l’activité de PPARγ. Par ailleurs, l’activation de ERK favorise la phosphorylation de facteurs de transcription, contrôlant positivement l’expression de PPARγ et de C/EBPα (CCAAT enhancer binding protein α) dans les cellules préadipocytaires afin d’induire leur différenciation [40]. Ainsi, l’activité de PPARγ devrait être finement modulée par la voie ERK dans certains types cellulaires, suggérant l’existence d’une régulation semblable pour la différenciation adéquate des trophoblastes.
Pour leur part, les mutants Lbp-1a-/- meurent à mi-gestation d’un sous-développement de la région du labyrinthe. Les vaisseaux embryonnaires ne pénètrent pas en profondeur la région du labyrinthe, ce qui s’accompagne d’une désorganisation vasculaire et d’une insuffisance placentaire. De plus, chez ces mutants, des structures semblables aux cellules MTG sont détectées. En revanche, l’expression des marqueurs de différenciation trophoblastique demeure intacte et les expériences d’agrégation avec des cellules tétraploïdes de type sauvage ne corrigent pas le phénotype placentaire des mutants Lbp-1a [32]. Donc, contrairement aux mutants associés à l’activation de la voie ERK/MAPK, les mutants Lbp-1a meurent à la suite d’un défaut primaire dans les tissus provenant de l’allantoïde. Les protéines LBP-1 sont phosphorylées par ERK, ce qui potentialise leur activité de liaison à l’ADN et leur activité transcriptionnelle [41, 42].
Conclusion
Le développement du labyrinthe dépend d’actions cellulaires non autonomes, telles que la production de facteurs paracrines embryonnaires sécrétés par l’allantoïde vers le chorion pour induire l’invagination des SynT-II dans la région du labyrinthe et permettre l’établissement de réseaux vasculaires fonctionnels [22]. Chez les mutants Mek1+/-Mek2+/-, la perte de vascularisation au niveau du placenta et la formation de cellules MTG seraient dues à l’absence de fonction de la kinase MEK1 dans les SynT-II [15]. Cependant, cette hypothèse provient d’expériences de délétion spécifique de Mek1 dans les SynT-II effectuées dans un environnement hétérozygote pour Mek2. Il ne peut donc être exclu que la perte d’un allèle de Mek2 dans l’allantoïde ou un autre tissu puisse contribuer au phénotype en influençant le niveau de phosphorylation de LBP-1a et son activité transcriptionnelle. Ainsi, on peut supposer que les protéines LBP-1 pourraient permettre la production de signaux par les tissus de l’allantoïde qui seraient intégrés par la voie ERK/MAPK dans les SynT-II. La voie ERK/MAPK jouerait donc un rôle dans différents types cellulaires du placenta autant pour réguler la production de signaux que pour leur intégration.
L’ensemble de ces données démontre l’importance de la voie ERK/MAPK dans l’organogenèse du placenta en participant à l’établissement de la région d’échanges materno-fœtaux, le labyrinthe. Cette région est composée de tissus d’origine extraembryonnaire (les trophoblastes) et embryonnaire (l’allantoïde). Une signalisation inadéquate de la voie ERK/MAPK dans les structures extraembryonnaires compromet la viabilité de l’embryon. Ainsi, la voie ERK/MAPK participe à la prolifération et la survie des trophoblastes ce qui permet la croissance de la région du labyrinthe. De plus, cette voie participe à la différenciation des SynT-II et, ainsi, à l’établissement de la barrière hématoplacentaire. Des études génétiques futures permettront d’élucider si la voie ERK/MAPK a un rôle dans d’autres types cellulaires trophoblastiques. De plus, les cibles cellulaires et les mécanismes contrôlés par cette voie restent encore à identifier.
Conflit d’intérêts
Les auteurs déclarent n’avoir aucun conflit d’intérêts concernant les données publiées dans cet article.
Remerciements
Les auteurs remercient les Drs Lucie Jeannotte et Olivier Boucherat pour leurs commentaires éclairés. Les travaux présentés bénéficient du support financier des instituts de recherche en santé du Canada (MOP-97801).
« On peut aussi obtenir le développement complet d’une souris indépendamment de la transmission germinale via des expériences dites de “complémentation d’embryons tétraploïdes”, ce qui représente le gold standard des tests de pluripotence. Cela consiste à agréger les cellules à tester avec des embryons tétraploïdes (obtenus par électrofusion d’embryons au stade 2 cellules), et à réimplanter l’agrégat dans l’utérus d’une femelle gravide. On peut aussi injecter les cellules à tester dans l’embryon tétraploïde après l’avoir cultivé jusqu’au stade blastocyste in vitro. A. Nagy a montré il y a vingt ans que les cellules embryonnaires tétraploïdes ne pouvaient pas (ou dans une très faible proportion et transitoirement) se développer en un embryon et contribuaient exclusivement aux annexes » (repris de [43]).
Références
- Rubinfeld H, Seger R. The ERK cascade: a prototype of MAPK signaling. Mol Biotechnol 2005 ; 31 : 151–174. [CrossRef] [PubMed] [Google Scholar]
- Wortzel I, Seger R. The ERK cascade: distinct functions within various subcellular organelles. Genes Cancer 2011 ; 2 : 195–209. [CrossRef] [PubMed] [Google Scholar]
- Fischer A, Katayama C, Pages G, et al. The role of erk1 and erk2 in multiple stages of T cell development. Immunity 2005 ; 23 : 431–443. [CrossRef] [PubMed] [Google Scholar]
- Ussar S, Voss T. MEK1 and MEK2, different regulators of the G1/S transition. J Biol Chem 2004 ; 279 : 43861–43869. [CrossRef] [PubMed] [Google Scholar]
- Orton RJ, Sturm OE, Vyshemirsky V, et al. Computational modelling of the receptor-tyrosine-kinase-activated MAPK pathway. Biochem J 2005 ; 392 : 249–261. [CrossRef] [PubMed] [Google Scholar]
- Shaul YD, Seger R. The MEK/ERK cascade: from signaling specificity to diverse functions. Biochim Biophys Acta 2007 ; 1773 : 1213–1226. [CrossRef] [PubMed] [Google Scholar]
- Rossant J, Cross J. Placental development: lessons from mouse mutants. Nat Rev Genet 2001 ; 2 : 538–548. [CrossRef] [PubMed] [Google Scholar]
- Dackor J, Strunk KE, Wehmeyer MM, Threadgill DW. Altered trophoblast proliferation is insufficient to account for placental dysfunction in Egfr null embryos. Placenta 2007 ; 28 : 1211–1218. [CrossRef] [PubMed] [Google Scholar]
- Hatano N, Mori Y, Oh-Hora M, et al. Essential role for ERK2 mitogen-activated protein kinase in placental development. Genes Cells 2003 ; 8 : 847–856. [CrossRef] [PubMed] [Google Scholar]
- Goldin SN, Papaioannou VE. Paracrine action of FGF4 during periimplantation development maintains trophectoderm and primitive endoderm. Genesis 2003 ; 36 : 40–47. [CrossRef] [PubMed] [Google Scholar]
- Schmidt C, Bladt F, Goedecke S, et al. Scatter factor/hepatocyte growth factor is essential for liver development. Nature 1995 ; 373 : 699–702. [CrossRef] [PubMed] [Google Scholar]
- Giroux S, Tremblay M, Bernard D, et al. Embryonic death of Mek1-deficient mice reveals a role for this kinase in angiogenesis in the labyrinthine region of the placenta. Curr Biol 1999 ; 9 : 369–372. [CrossRef] [PubMed] [Google Scholar]
- Bissonauth V, Roy S, Gravel M, et al. Requirement for Map2k1 (Mek1) in extra-embryonic ectoderm during placentogenesis. Development 2006 ; 133 : 3429–3440. [CrossRef] [PubMed] [Google Scholar]
- Catalanotti F, Reyes G, Jesenberger V, et al. A Mek1-Mek2 heterodimer determines the strength and duration of the Erk signal. Nat Struct Mol Biol 2009 ; 16 : 294–303. [CrossRef] [PubMed] [Google Scholar]
- Nadeau V, Guillemette S, Bélanger L-F, et al. Map2k1 and Map2k2 genes contribute to the normal development of syncytiotrophoblasts during placentation. Development 2009 ; 136 : 1363–1374. [CrossRef] [PubMed] [Google Scholar]
- Sachs M, Brohmann H, Zechner D, et al. Essential role of Gab1 for signaling by the c-Met receptor in vivo. J Cell Biol 2000 ; 150 : 1375–1384. [CrossRef] [PubMed] [Google Scholar]
- Galabova-Kovacs G, Matzen D, Piazzolla D, et al. Essential role of B-Raf in ERK activation during extraembryonic development. Proc Nat Acad Sci USA 2006 ; 103 : 1325–1330. [CrossRef] [Google Scholar]
- Huser M, Luckett J, Chiloeches A, et al. MEK kinase activity is not necessary for Raf-1 function. EMBO J 2001 ; 20 : 1940–1951. [CrossRef] [PubMed] [Google Scholar]
- Shaw AT, Meissner A, Dowdle JA, et al. Sprouty-2 regulates oncogenic K-ras in lung development and tumorigenesis. Genes Dev 2007 ; 21 : 694–707. [CrossRef] [PubMed] [Google Scholar]
- Watson ED, Cross JC. Development of structures and transport functions in the mouse placenta. Physiology (Bethesda) 2005 ; 20 : 180–193. [CrossRef] [PubMed] [Google Scholar]
- Cross JC, Nakano H, Natale DRC, et al. Branching morphogenesis during development of placental villi. Differentiation 2006 ; 74 : 393–401. [CrossRef] [PubMed] [Google Scholar]
- Cross JC. How to make a placenta: mechanisms of trophoblast cell differentiation in mice: a review. Placenta 2005 ; 26 (suppl A) : S3–S9. [CrossRef] [PubMed] [Google Scholar]
- Cross J. Genetic insights into trophoblast differentiation and placental morphogenesis. Semin Cell Dev Biol 2000 ; 11 : 105–113. [CrossRef] [PubMed] [Google Scholar]
- Nagai A, Takebe K, Nio-Kobayashi J, et al. Cellular expression of the monocarboxylate transporter (MCT) family in the placenta of mice. Placenta 2010 ; 31 : 126–133. [CrossRef] [PubMed] [Google Scholar]
- Simmons DG, Natale DRC, Begay V, et al. Early patterning of the chorion leads to the trilaminar trophoblast cell structure in the placental labyrinth. Development 2008 ; 135 : 2083–2091. [CrossRef] [PubMed] [Google Scholar]
- Charron J, Jeannotte L. Le rôle essentiel de MEK1 lors de l’angiogenèse placentaire. Med Sci (Paris) 1999 ; 15 : 1155–1157. [CrossRef] [Google Scholar]
- Gee E, Milkiewicz M, Haas TL. p38 MAPK activity is stimulated by vascular endothelial growth factor receptor 2 activation and is essential for shear stress-induced angiogenesis. J Cell Physiol 2010 ; 222 : 120–126. [CrossRef] [PubMed] [Google Scholar]
- Bélanger LF, Roy S, Tremblay M, et al. Mek2 is dispensable for mouse growth and development. Mol Cell Biol 2003 ; 23 : 4778–4787. [CrossRef] [PubMed] [Google Scholar]
- Fernandez-Serra M, Consales C, Livigni A, Arnone MI. Role of the ERK-mediated signaling pathway in mesenchyme formation and differentiation in the sea urchin embryo. Dev Biol 2004 ; 268 : 384–402. [CrossRef] [PubMed] [Google Scholar]
- Qian X, Esteban L, Vass W, et al. The Sos1 and Sos2 Ras-specific exchange factors: differences in placental expression and signaling properties. EMBO J 2000 ; 19 : 642–654. [CrossRef] [PubMed] [Google Scholar]
- Parast MM, Yu H, Ciric A, et al. PPARgamma regulates trophoblast proliferation, promotes labyrinthine trilineage differentiation. PLoS One 2009 ; 4 : e8055. [CrossRef] [PubMed] [Google Scholar]
- Parekh V, McEwen A, Barbour V, et al. Defective extraembryonic angiogenesis in mice lacking LBP-1a, a member of the grainyhead family of transcription factors. Mol Cell Biol 2004 ; 24 : 7113–7129. [CrossRef] [PubMed] [Google Scholar]
- Barak Y, Nelson MC, Ong ES, et al. PPAR gamma is required for placental, cardiac, and adipose tissue development. Mol Cell 1999 ; 4 : 585–595. [CrossRef] [PubMed] [Google Scholar]
- Burgermeister E, Chuderland D, Hanoch T, et al. Interaction with MEK causes nuclear export and downregulation of peroxisome proliferator-activated receptor gamma. Mol Cell Biol 2007 ; 27 : 803–817. [CrossRef] [PubMed] [Google Scholar]
- Froment P, Gizard F, Staels B, et al. Un rôle pour PPARγ dans la reproduction ? Med Sci (Paris) 2005 ; 21 : 507–511. [CrossRef] [EDP Sciences] [PubMed] [Google Scholar]
- Kubota N, Terauchi Y, Miki H, et al. PPAR gamma mediates high-fat diet-induced adipocyte hypertrophy and insulin resistance. Mol Cell 1999 ; 4 : 597–609. [CrossRef] [PubMed] [Google Scholar]
- Cross JC, Anson-Cartwright L, Scott IC. Transcription factors underlying the development and endocrine functions of the placenta. Recent Prog Horm Res 2002 ; 57 : 221–234. [CrossRef] [PubMed] [Google Scholar]
- Shalom-Barak T, Nicholas JM, Wang Y, et al. Peroxisome proliferator-activated receptor gamma controls Muc1 transcription in trophoblasts. Mol Cell Biol 2004 ; 24 : 10661–10669. [CrossRef] [PubMed] [Google Scholar]
- Camp HS, Tafuri SR. Regulation of peroxisome proliferator-activated receptor gamma activity by mitogen-activated protein kinase. J Biol Chem 1997 ; 272 : 10811–10816. [CrossRef] [PubMed] [Google Scholar]
- Prusty D. Activation of MEK/ERK signaling promotes adipogenesis by enhancing peroxisome proliferator-activated receptor gamma (PPARgamma) and C/EBPalpha gene expression during the differentiation of 3T3–L1 preadipocytes. J Biol Chem 2002 ; 277 : 46226–46232. [CrossRef] [PubMed] [Google Scholar]
- Pagon Z, Volker J, Cooper GM, Hansen U. Mammalian transcription factor LSF is a target of ERK signaling. J Cell Biochem 2003 ; 89 : 733–746. [CrossRef] [PubMed] [Google Scholar]
- Volker JL, Rameh LE, Zhu Q, et al. Mitogenic stimulation of resting T cells causes rapid phosphorylation of the transcription factor LSF and increased DNA-binding activity. Genes Dev 1997 ; 11 : 1435–1446. [CrossRef] [PubMed] [Google Scholar]
- Coulombel L. Pluripotence : une définition à géométrie variable. Med Sci (Paris) 2009 ; 25 : 798–801. [CrossRef] [EDP Sciences] [PubMed] [Google Scholar]
Liste des tableaux
Phénotype placentaire des souris mutantes pour des protéines impliquées dans la voie ERK/MAPK.
Liste des figures
|
Figure 1. Structure du placenta de la souris : organisation des différentes couches cellulaires de la région du labyrinthe de souris. A. Représentation schématique du placenta de souris (panneau de gauche) et coupe histologique d’un placenta d’un embryon au 12,5e jour de gestation (panneau de droite). La ligne pointillée sépare les parties maternelle et embryonnaire. B. Schéma de l’organisation de la triple couche de trophoblastes et de l’endothélium fœtal dans la région du labyrinthe permettant la formation de la barrière hématoplacentaire (panneau de gauche). Visualisation de la double couche de syncytiotrophoblastes par immunofluorescence avec des anticorps contre les transporteurs de monocarboxylate MCT1 (en rouge) et MCT4 (en vert) (panneau de droite) [24]. MCT1 et MCT4 sont utilisés comme marqueurs spécifiques de la membrane apicale des SynT-I et de la membrane basale des SynT-II, respectivement. Dans cette expérience, les noyaux sont marqués en bleu par une coloration au DAPI. Les érythrocytes maternels sont détectés en orange (du fait de l’autofluorescence), alors que les érythrocytes fœtaux ont un noyau bleu et un cytoplasme verdâtre. Les érythrocytes et les sinus maternels sont délimités par la couche de SynT-I (en rouge), alors que les érythrocytes et les cellules endothéliales des vaisseaux sanguins fœtaux sont inclus à l’intérieur d’une couche de SynT-II (en vert). Dans le vaisseau sanguin fœtal indiqué (VSF), une cellule endothéliale (CE) accolée sur la partie interne de la couche de SynT-II définie par le marquage MCT4 est visible. Les circulations sanguines fœtale et maternelle sont séparées par trois couches étanches constituées des cellules endothéliales vasculaires fœtales, des SynT-I et des SynT-II. CE, cellule endothéliale ; EF, érythrocyte fœtal ; EM, érythrocyte maternel ; SM, sinus maternel ; SynT-I, syncytiotrophoblaste de type I ; SynT-II, syncytiotrophoblaste de type II ; VSF, vaisseau sanguin fœtal. |
| Dans le texte | |
|
Figure 2. Activation différentielle des voies ERK et p38 MAPK dans les cellules du labyrinthe. Le réseau vasculaire embryonnaire est révélé à l’aide d’un anticorps anti-CD31 (cluster of differentiation 31) alors que l’activation des voies de signalisation ERK/MAPK et p38/MAPK est mise en évidence avec des anticorps contre les formes phosphorylées de ERK (p-ERK) et de p38 (p-p38), respectivement. Les images sont prises en contraste interférentiel. Les érythrocytes embryonnaires et maternels sont détectables dans les vaisseaux fœtaux (VF) et les sinus maternels (SM), respectivement. Les érythrocytes embryonnaires sont plus gros et nucléés. L’activation de la voie p38/MAPK est restreinte aux vaisseaux sanguins fœtaux alors que l’activation de ERK est localisée en périphérie des sinus maternels. Les flèches noires indiquent les cellules endothéliales vasculaires fœtales et les flèches blanches les trophoblastes bordant les sinus maternels. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.