")
")
| Issue |
Med Sci (Paris)
Volume 27, Number 11, Novembre 2011
Imagerie et cognition
|
|
|---|---|---|
| Page(s) | 1000 - 1008 | |
| Section | M/S Revues | |
| DOI | https://doi.org/10.1051/medsci/201127111000 | |
| Published online | 30 novembre 2011 | |
Apport des « neurosciences de la décision » à l’étude des comportements alimentaires et de l’obésité
Imagerie et cognition (10)
Food behaviour and obesity: insights from decision neuroscience
1
Laboratoire de psychologie cognitive (UMR 6146), Université de Provence et CNRS, Aix-Marseille Université, Pôle 3C-Case D, 3, place Victor Hugo, 13331 Marseille Cedex 3, France
2
Institut d’administration des entreprises (IAE-CERGAM), Université Paul Cézanne, Aix-Marseille Université, Aix-en-Provence, France
3
Centre de recherche en économie et en management (UMR 6211), IGR-IAE de Rennes, Université de Rennes 1, Rennes, France
4
Laboratoire de neurosciences de la décision, Département de marketing, INSEAD, Fontainebleau, France
5
Laboratoire de neurosciences cognitives (INSERM U960), École normale supérieure et École des neurosciences de Paris, Paris, France
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Résumé
Les « neurosciences de la décision » étudient l’activité cérébrale associée aux choix. Elles apportent de fait un éclairage nouveau sur les mécanismes comportementaux et cérébraux qui les sous-tendent. Ce champ d’investigation permet de renouveler le questionnement de la « théorie du choix rationnel » issue des sciences économiques. Cette dernière repose sur une rationalité idéalisée postulant qu’après avoir évalué toutes les possibilités offertes et notamment le rapport coût/gain, les individus choisissent toujours la meilleure alternative, c’est-à-dire celle qui maximise leur utilité. La neuro-imagerie fonctionnelle pointe les changements d’activité cérébrale associés à la prise de décision, en mettant en évidence l’intervention d’un réseau cérébral (ou circuit) dit « de la récompense » dans l’évaluation des objectifs, la motivation à agir et les choix comportementaux. La compréhension de l’intervention et du rôle de ce réseau dans la prise de décision apporte des éclairages nouveaux dans de nombreux domaines, et notamment ceux des comportements de consommation. De tels résultats dans le cadre d’études des comportements (et des désordres) alimentaires devraient à présent contribuer à l’élaboration de campagnes de prévention en santé publique, plus adaptées à la réalité du comportement hédonique des consommateurs, loin de l’idéal économique rationnel.
Abstract
Neuroimaging allows to estimate brain activity when individuals are doing something. The location and intensity of this estimated activity provides information on the dynamics and processes that guide choice behaviour and associated actions that should be considered a complement to behavioural studies. Decision neuroscience therefore sheds new light on whether the brain evaluates and compares alternatives when decisions are made, or if other processes are at stake. This work helped to demonstrate that the situations faced by individuals (risky, uncertain, delayed in time) do not all have the same (behavioural) complexity, and are not underlined by activity in the cerebral networks. Taking into account brain dynamics of people (suffering from obesity or not) when making food consumption decisions might allow for improved strategies in public health prevention, far from the rational choice theory promoted by neoclassical economics.
© 2011 médecine/sciences – Inserm / SRMS
Imaginez-vous devant un distributeur de nourriture. Pour 1€ vous avez la possibilité d’acheter une barre chocolatée ou une pomme. Quels facteurs vont déterminer votre choix ? Vous pouvez vous dire que la pomme a l’air juteuse, qu’elle doit être bonne pour la santé. La barre chocolatée certes est plus calorique, mais vous adorez le chocolat, et la manger vous procurera assurément plus de plaisir que la pomme. Mais ce n’est pas tout. Comme le suggère une expression plus que millénaire : « On mange avec les yeux ». En effet, le simple fait d’observer ces deux aliments déclenche en vous, sans le savoir, c’est-à-dire de manière non consciente, une représentation de leur goût plaisant. Et ceci sans compter l’éventuelle influence que la publicité et/ou certains de vos proches peuvent avoir sur ces mêmes goûts et sur vos choix de consommation. L’un dans l’autre, comme pour nombre d’autres comportements, ceux liés à notre alimentation résultent d’interactions complexes entre les dynamiques comportementales et cérébrales, individuelles et sociales.
L’objectif de cet article est dès lors de présenter, à travers quelques études jugées pertinentes, les apports de l’imagerie cérébrale fonctionnelle à la compréhension des mécanismes de décision. Plus spécifiquement, nous nous intéresserons à des comportements alimentaires qui bien souvent ne sont pas les meilleurs d’un point de vue sanitaire. Ils ont toutefois une caractéristique indéniable : même s’ils sont mauvais pour notre santé et notre bien-être, nombre d’entre nous faisons le choix de consommer des aliments gras et sucrés - le plus souvent en pleine connaissance de leurs effets délétères à plus ou moins court terme. Prendre une telle décision revient dès lors, d’un point de vue économique, à faire un choix sous optimal.
Dans les sections qui suivent, nous présenterons tout d’abord des investigations cérébrales portant sur la vision des aliments. Puis nous essaierons d’apporter un éclairage à la question du calcul coût/avantage qui, pour certains, accompagnerait nos décisions, car (fort heureusement ?) les êtres humains sont loin d’être des « agents rationnels » comme encore trop d’économistes aiment à les penser. Ensuite, nous présenterons des travaux récents portant sur la décision d’achats alimentaires, notamment ceux qui permettent de comparer les différences de traitement entre les personnes souffrant d’obésité et des populations contrôles.
Neurosciences et perception visuelle des aliments
De nombreux travaux utilisant la technique de l’imagerie par résonance magnétique fonctionnelle (IRMf) se sont concentrés ces dernières années sur la perception du goût évoquée par la seule vue d’un aliment.
Une revue de la littérature, récemment publiée à partir de ces études, nous permet de dresser trois conclusions principales [1]. Tout d’abord - et en recoupant cette revue avec un article plus ancien [2] -, à la vue d’un aliment, il apparaît manifestement que ce dernier déclenche en nous de l’activité cérébrale dans des régions reflétant des « inférences gustatives ». Un réseau cérébral révèle une activité significativement plus élevée lorsque les activations liées à la vision d’un produit alimentaire sont contrastées avec celles d’un produit non alimentaire. Il s’agirait principalement des aires suivantes : le gyrus fusiforme postérieur (gauche et droit), associé à la reconnaissance des objets, le cortex orbitofrontal latéral gauche, corrélé au plaisir subjectif, et le cortex insulaire (médian), plus traditionnellement relié à l’aliment (Figure 1A ; rouge) [1].
 |
Figure 1. Méta-analyse d’études sur la vision de nourriture utilisant l’imagerie par résonance magnétique fonctionnelle. A. Activité significativement plus élevée lorsque l’individu voit de la nourriture en comparaison d’objets non alimentaires (rouge). B. Activité significativement plus élevée lorsque l’individu voit de la nourriture en état de faim versus en état de satiété (bleu), et activité significativement plus élevée lorsque la nourriture vue est plus calorique que « diététique » (moins calorique ; vert) (adapté de [1]). |
Ensuite, les régions cérébrales participant à ce réseau, et l’intensité de leurs activités, varient avec l’état de l’individu ou la nature des aliments présentés. Ainsi, lorsque l’individu a faim, on observe, par contraste avec l’état de satiété, des activations amygdaliennes et proches du gyrus parahippocampique droit, associées au traitement de la récompense, en plus de celles situées dans le cortex orbitofrontal latéral gauche (Figure 1B ; bleu). Ou encore, les activations que provoque la vision de produits gras (par exemple : glaces, cookies, burgers, etc.), comparées à celles que suscite la présentation de produits moins caloriques (salades, fruits, poissons, légumes, etc.), sont situées principalement dans l’hypothalamus (notamment impliqué dans la régulation de la prise alimentaire et de la glycémie) et dans le striatum ventral (Figure 1C ; vert).
Enfin, il ressort de l’examen des résultats de ces expérimentations que les activations rapportées dans la méta-analyse sont très variables d’un protocole à l’autre. Il convient donc de se garder de livrer des conclusions définitives sur un réseau cérébral qui serait « propre à » la vue d’un aliment en particulier. Par exemple, dans le cas des activations associées à la vue d’un aliment (par contraste avec celles associées à la vue d’un produit non alimentaire), les quatre régions indiquées ne sont présentes que dans à peine plus d’un tiers des études [1] (Encadré 1).
Théorie économique de la décision
Maintenant que vous avez vu les produits alimentaires qui s’offrent à vous, comment votre choix va-t-il s’arrêter sur la pomme ou la barre chocolatée1 ?
Première piste, longtemps suggérée dans toute une partie de la littérature en sciences économiques : ce serait suite à un calcul coût/avantage des différentes options qui se présentent à vous que vous choisiriez un aliment plutôt qu’un autre. En d’autres termes, qu’ai-je à gagner et qu’ai-je à perdre ? Cette vision très « computationnelle » et rationnelle de la prise de décision est l’héritage direct de la théorie économique néo-classique (Encadré 2).
1 Variabilité et représentativité des résultats
Des précautions quant à la représentativité des résultats doivent être prises pour toute étude expérimentale, qu’elle implique ou non la neuro-imagerie, l’aliment ou non - et il faudra assurément redoubler de prudence sur l’aliment au regard de la grande variabilité culturelle qui préside à nos pratiques alimentaires. Deux autres revues récentes de la littérature sont sans équivoque à ce sujet. D’une part, d’une étude à l’autre, lorsque les études expérimentales portent sur des sujets occidentaux, il faut garder à l’esprit que ces derniers font plus figure d’exception que d’individus représentatifs de la population globale, qu’il s’agisse de tâches de perception, de catégorisation ou de jugement [3]. D’autre part, les résultats des études expérimentales ne sont pas nécessairement robustes, c’est-à-dire facilement reproductibles. Une publication récente [4] a effectivement montré que la reproductibilité des résultats d’expérimentations en IRMf n’excédait pas 50 %, même dans les cas où les mêmes tâches et les mêmes stimulus étaient présentés au même groupe de participants !1
2 Principaux postulats de la théorie du choix rationnel
La théorie économique postule que l’homme économique (ou homo oeconomicus) est un être rationnel piloté par son intérêt bien compris [5–7]. Il est donc fait l’hypothèse que les agents économiques ont des objectifs cohérents entre eux, une connaissance appropriée de leur environnement et ont des préférences stables dans le temps. Dans une telle perspective, les décideurs sont des « agents » capables de faire une analyse coût/avantage pour déterminer l’utilité de chaque option en traitant toutes les informations disponibles et en estimant correctement les probabilités qui leurs sont associées. En simplifiant, l’utilité est une forme de mesure du bien-être attendu pour chaque option qui s’offre à un individu. Dès lors, une décision serait théoriquement prise à partir de la comparaison de l’utilité de chaque alternative. Il est considéré que les besoins des agents sont séparables, ce qui permet d’additionner les utilités. L’exigence de comparabilité qui en découle nécessite l’introduction d’un étalon de mesure servant d’intermédiaire. C’est le plus souvent la monnaie qui joue ce rôle. Les besoins sont alors satisfaits par l’obtention des biens et des services désirés, à partir de l’optimisation de la fonction d’utilité (objectif) sur la base du revenu et des prix supposés donnés (contraintes).
Mieux, cette théorie repose sur la croyance que nous choisissons toujours ce que nous préférons, faisant ainsi de nos choix les révélateurs de nos préférences [8].
Mais est-ce véritablement aussi simple ? Une personne choisit-elle toujours l’option qu’elle préfère après avoir pesé le pour et le contre lorsqu’elle doit prendre une décision ? Pas vraiment. Pourquoi ? Tout simplement parce que cette personne est humaine et que ses décisions sont le fruit d’une tension permanente entre sa rationalité et ses émotions, modulée par ses buts personnels et par son contexte physique et social [9]. De nombreux travaux de psychologie sociale montrent par exemple que nos « choix », alimentaires ou autres, résultent en réalité assez souvent de conduites socialement contraintes mais rationalisées après coup, d’où le sentiment d’une décision librement consentie. Dans ce cas la rationalité suit le comportement plus qu’elle ne le précède, en totale opposition avec le modèle économique standard [10]. D’autres travaux montrent que les individus peuvent ne pas avoir conscience des processus qu’ils mettent en œuvre pour l’atteinte efficace d’une utilité, et en effet attribuer l’atteinte du but à une motivation monétaire objectivement inefficace [11].
Face à une représentation de la prise de décision de fait simpliste car décontextualisée - mais bien pratique pour développer des modèles mathématiques - plusieurs auteurs se sont attelés à mettre en évidence les capacités cognitives limitées de l’(hypothétique) agent économique [12–14]. Afin d’alléger leurs efforts cognitifs en situation de résolution de problèmes, les individus ont généralement recours à des heuristiques leur permettant de simplifier la représentation des problèmes à résoudre. Celles-ci entraînent cependant des biais dans la comparaison des options, modifiant les processus d’apprentissage et réduisant la recherche d’informations [15–18].
L’imagerie fonctionnelle permet d’analyser certaines des étapes qui composeraient la prise de décision, dont le possible recours aux heuristiques. Cette méthodologie offre une estimation de l’activité cérébrale au moment de l’évaluation des options, dans le traitement des probabilités et lors de la prise de décision.
Dopamine, récompense et consommation
Certaines recherches en neurosciences de la décision ont eu pour but d’identifier les régions cérébrales impliquées théoriquement dans le calcul de l’utilité et les mécanismes par lesquels le cerveau est supposé évaluer et comparer les récompenses associées à chaque alternative [19, 20]. L’hypothèse d’un tel fonctionnement, discutable, et discutée, reste largement répandue. Elle représente même le courant dominant en neuroéconomie [21]2 (voir toutefois [22] pour une proposition alternative).
Dans une telle perspective, pour que les alternatives soient comparables, il est postulé qu’elles doivent être transformées par le cerveau en une même unité de mesure. De nombreux auteurs font l’hypothèse qu’un neurotransmetteur, la dopamine, pourrait jouer un rôle similaire à celui de l’utilité dans la sélection d’objectifs [23]. Des neurones dopaminergiques ont été trouvés dans plusieurs parties du cerveau, permettant d’identifier un circuit de la récompense (Figure 2). Ainsi, lors d’une prise de décision, la dopamine jouerait un rôle dans l’évaluation des options, le calcul des probabilités et la motivation à agir [23].
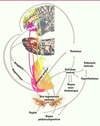 |
Figure 2. Schéma des principales structures et connexions composant le circuit de la récompense. Flèches rouges : efférences du cortex préfrontal ventromédian (vmPFC) vers le striatum ventral ; flèches oranges : efférences de la partie dorsale du cortex cingulaire antérieur (dACC) vers le striatum ventral ; flèches jaunes : efférences du cortex préfrontal dorsal (dPFC) vers le striatum ventral ; flèches marrons : principales connexions du circuit de la récompense (adapté de [23]). |
Plusieurs travaux impliquant la technique de l’IRMf ont mis en évidence l’intervention du système de la récompense dans la prise en compte de la valeur appétitive de la nourriture [24], des drogues [25], ou encore des gains monétaires [26]. Si les neurones dopaminergiques sont initialement activés par l’arrivée de la récompense, leur rôle dans l’apprentissage les fait ensuite réagir directement à la répétition des stimulus conditionnés annonçant la récompense [27], comme cela peut devenir le cas lorsqu’une publicité pour un produit alimentaire est vue plusieurs fois.
Néanmoins, si l’on se place dans la perspective de la théorie du choix rationnel, les individus devraient toujours choisir le bien qui, pour eux, a la plus grande valeur. Dans le cadre du choix alimentaire, la valeur n’est pas uniquement déterminée par les propriétés physiques de la nourriture. Cette valeur est déterminée par le plaisir instantané (le goût ressenti au moment de la consommation), l’anticipation et l’apprentissage (connaissance du produit) et la motivation à obtenir le produit basée sur le plaisir attendu de la consommation. Toujours dans une perspective rationnelle, si lors d’un premier achat un consommateur venait à acheter un produit dans des quantités supérieures à la satisfaction de ses besoins, lors d’un second achat il devrait normalement rectifier son erreur d’estimation [28]. Une telle hypothèse repose sur la croyance que les individus sont capables de faire coïncider le plaisir attendu de la consommation et le plaisir au moment de la consommation.
Une découverte majeure en neurosciences de la décision conduit à une remise en cause de ce postulat, en mettant en évidence l’existence d’une dissociation dans le traitement cérébral entre le plaisir instantané et le plaisir attendu [29]. Dès lors, il n’est pas certain que l’apprentissage puisse faire coïncider le plaisir attendu avec le plaisir instantané et, par là même, que les choix permettent de révéler les préférences du consommateur. Cette dissociation peut conduire le consommateur à persister dans son comportement et à ne pas modifier sa décision d’achat malgré son expérience.
Ainsi, des études ont-elles indiqué que le niveau de dopamine était davantage corrélé avec le plaisir attendu de la consommation qu’avec le plaisir instantané [30]. Des expériences menées sur des rats [31], puis élargies à des fumeurs [32] ont indiqué que lorsque l’on perturbe à la baisse la production de dopamine, les sujets présentent une plus faible motivation à obtenir une récompense (sucre, cigarette) mais ressentent un plaisir identique lors de la consommation. Dans le sens inverse, des sujets parkinsoniens chez qui la production de dopamine est augmentée ont un plaisir identique au moment de la consommation (médicaments), mais leur motivation augmente [33] et les récompenses potentielles sont surestimées [34].
Ces travaux - que l’on retrouve dans la littérature anglosaxonne sous la dichotomie « liking-wanting » - amènent ainsi à réduire le rôle du plaisir instantané pourtant au cœur des modèles économiques rationnels (dans la valeur allouée au produit au moment de la prise de décision). Ceci est confirmé par de récents travaux montrant que lorsque des sujets consomment des aliments appréciés jusqu’à satiété, la motivation peut demeurer inchangée voire augmenter alors que la faim et l’appréciation de la nourriture sont réduites [35]. D’où un risque de surconsommation puisque la motivation à obtenir un aliment peut avoir une certaine indépendance vis-à-vis des processus homéostatiques liés aux besoins métaboliques.
Contexte, effort cognitif et choix intertemporel
Les situations auxquelles doivent faire face les individus n’ont pas toutes la même complexité [5, 6]. Celles risquées et incertaines sont généralement perçues comme plus complexes et les individus tendent à les éviter [5, 6]. De plus, il a été montré que l’aversion au risque varie selon que les options proposées à un individu sont présentées sous forme de gains ou de pertes [36]. L’utilisation de l’IRMf a permis de mieux comprendre pourquoi une telle variation existe. Dans une étude, des sujets devaient choisir entre plusieurs loteries (risquée, non risquée et incertaine), auxquelles étaient attachées des probabilités et des paiements présentés sous forme de gains ou de pertes (Figure 3) [37].
 |
Figure 3. Risque et incertitude. La tâche du participant est de choisir entre deux paris en fonction des distributions de probabilités et de revenus pour chacune des billes constituant l’urne. Les chiffres au-dessus des flèches indiquent la distribution des billes rouges, noires et jaunes dans les urnes. Les chiffres en dessous des flèches indiquent les gains en dollars. Deux conditions sont présentées : A. Les sujets doivent choisir entre deux paris risqués sous forme de gain. Le pari sur la gauche a une variance plus faible et est considéré comme moins risqué. B. Les sujets doivent choisir entre un pari présentant une part d’incertitude (à gauche) et le même pari risqué que la condition précédente en A (à droite). Les flèches obliques indiquent qu’il y a un total de 60 billes noires et jaunes dans le pari 1. Le nombre précis de billes noires et vertes n’est pas connu (adapté de [37]). |
L’analyse comportementale indique que les sujets évitent le risque lorsque les loteries sont présentées sous forme de gains, alors qu’en présence de loteries présentées sous forme de pertes, l’aversion au risque n’est pas avérée et les participants modifient leurs préférences. L’analyse révèle également que, contrairement au risque, l’incertitude n’est préférée dans aucune des conditions.
L’observation de l’activité cérébrale apporte des précisions indiquant des traitements différents. Ainsi, en situation d’incertitude, contrairement aux autres situations, on n’observe aucun changement significatif d’activité dans la partie ventrale du cortex préfrontal [37]. Cette partie du cerveau est notamment associée à l’attente de récompenses [38]. Une hypothèse serait qu’en situation d’incertitude, les individus n’auraient pas assez de ressources cognitives pour évaluer les récompenses futures. Concernant les autres conditions, un réseau (dorso-médian) interviendrait uniquement dans le traitement des pertes en situation de risque et un autre réseau (ventro-médian) interviendrait dans le traitement des autres situations (gain non risqué, perte non risquée, gain risqué). En comparaison du réseau ventro-médian, le dorso-médian est généralement associé à une plus grande mobilisation de ressources cognitives et computationnelles. Une telle observation au niveau cérébral est cohérente avec les résultats comportementaux. En effet, ces derniers indiquent des erreurs d’estimation moins importantes en situation de pertes risquées.
Les choix intertemporels sont également des situations susceptibles de poser problème aux individus. Il s’agit de décisions qui impliquent des arbitrages entre des coûts et des bénéfices survenant à différents moments (futur proche, éloigné, etc.). Ces choix découlent généralement d’un conflit entre une préférence impulsive pour les gains présents et une prise en compte plus rationnelle des gains futurs. Certains auteurs considèrent que nous ne sommes pas capables d’appréhender parfaitement le temps, ce qui conduirait à surévaluer les récompenses présentes et à sous-évaluer les récompenses futures [39]. L’IRMf a été utilisée pour approcher l’activité cérébrale au moment où les individus effectuent des choix intertemporels [40]. Dans cette étude, les sujets doivent faire des choix impliquant des récompenses immédiates et des récompenses supérieures mais décalées dans le temps. Une activité importante dans les aires associées au réseau de la récompense est observée lorsque les sujets choisissent les récompenses immédiates. En revanche, d’autres régions (cortex préfrontal dorso-latéral, cortex pariétal postérieur), interviennent quand le choix se porte sur des options décalées dans le temps. Ces régions sont généralement associées avec le calcul, ou encore la résolution de problèmes et le raisonnement [41], suggérant que le choix de ces options demande plus d’effort cognitif voire plus de contrôle. Pour les auteurs, deux réseaux interviendraient dans le traitement de ces choix, un réseau lié à l’impulsivité, plus actif pour évaluer les récompenses actuelles, et un réseau de contrôle plus actif pour évaluer les récompenses à venir. Ces deux réseaux seraient présents chez tous les individus, leurs interactions permettraient d’arrêter les choix.
Sélection de la nourriture, plaisir ressenti et décision d’achat
La décision alimentaire - comme l’ensemble des décisions - reposerait sur la valorisation des différentes options. Dans le cadre de la décision d’achat, la volonté de payer (montant maximum que l’acheteur est prêt à donner pour obtenir le produit) constitue une composante essentielle de la transaction et un indicateur de la valorisation du produit. Mais pas uniquement. Avec l’acquisition en IRMf de données cérébrales au moment de la décision d’achat, il serait possible de mieux comprendre comment le cerveau effectue des calculs dans les transactions quotidiennes (si c’est bien ainsi qu’il procède) [42]. Des « junk food » (barre chocolatée, chips) sont présentées à des sujets qui n’ont pas mangé depuis plusieurs heures. Des dollars leur sont distribués en leur expliquant qu’ils peuvent soit garder l’argent, soit l’utiliser pour acheter les aliments au cours de l’expérience. Il leur est demandé d’indiquer combien ils sont prêts à payer pour chaque aliment (0, 1, 2 ou 3 $). Ce montant est considéré comme une mesure comportementale de l’utilité des aliments. Il apparaît qu’au moment où un hypothétique calcul d’utilité s’opère, l’activité du cerveau augmente de manière significative dans le cortex orbito-frontal médian et le cortex préfrontal dorso-latéral [42]. Les résultats montrent aussi que ces activités varient positivement avec l’importance du montant que les sujets sont prêts à investir.
Nous avons indiqué précédemment que les gains et les pertes (monétaires) n’étaient pas traités de la même manière dans le cerveau, notamment que les pertes risquées demandent plus d’efforts cognitifs [37]. Dans une autre étude [43], l’expérience décrite ci-dessus a été renouvelée en comparant les sollicitations cérébrales au moment du calcul de l’utilité pour des aliments appétissants ou rebutants, afin de voir si deux réseaux différents intervenaient. Dans cette expérience les sujets pouvaient par ailleurs attribuer des montants négatifs aux aliments (– 1, – 2, – 3 $). Les résultats indiquent que l’activité du cortex orbito-frontal et celle du cortex préfrontal dorso-latéral sont aussi bien corrélées avec l’évaluation positive des aliments qu’avec leur évaluation négative.
Si l’obésité résulte d’une multitude de facteurs aussi bien physiologiques, socio-psychologiques que socio-économiques, l’étude du cerveau dans le cadre de la récompense alimentaire s’est récemment révélée porteuse d’informations concernant la compréhension de la décision de consommation et des troubles alimentaires qui vont à rebours de la théorie rationnelle.
En effet, il est postulé dans cette théorie de la décision que les choix révèlent les préférences et ainsi que les aliments plaisants sont ceux choisis par les individus et, de fait, sont également ceux qui déclenchent une libération importante de dopamine dans le réseau de la récompense. Cependant si le plaisir durant la consommation et le plaisir attendu de la consommation sont liés, ils n’activent pas nécessairement de la même manière le réseau de la récompense [44]. L’IRMf met au jour les différences de sollicitations cérébrales et la possibilité d’une envie de manger sans plaisir au moment de la consommation [45].
Certaines études ont indiqué que les personnes souffrant d’obésité disposent d’un niveau de récepteurs dopaminergiques plus réduit, diminuant le plaisir ressenti durant la consommation d’aliments [45, 46]. Le plaisir étant réduit, les individus avec moins de récepteurs dopaminergiques mangeraient moins, ou seraient indifférents à la consommation de la pomme et de la barre chocolatée. Il semble au contraire que les personnes souffrant d’obésité consomment plus afin d’obtenir le même niveau de plaisir qu’une personne ne souffrant pas d’obésité. De plus, cette réduction de plaisir ne les rendrait pas indifférents aux aliments consommés. Il ressort d’une étude en IRMf que si les personnes obèses ont une plus faible activité au niveau du noyau caudé au cours de la consommation de milk-shake, ils présentent aussi une plus forte activité au niveau du cortex gustatif et somato-sensoriel (codant les aspects sensoriels et hédoniques des aliments) en réponse à l’apport prévu de milk-shake [45]. Ainsi deux réseaux, relativement indépendants, interviendraient dans l’évaluation de la nourriture, et la réduction du plaisir au moment de la consommation ne réduirait pas nécessairement le plaisir attendu de la consommation [44]. Comme dans l’addiction, la prise alimentaire peut semble-t-il devenir irrésistible alors même que la récompense (décharge dopaminergique) diminue [46]. Ceci va dans le sens d’études menées sur les choix intertemporels monétaires, indiquant que les personnes souffrant d’obésité choisissent davantage les options présentes plutôt que les gains décalés dans le temps, en comparaison de sujets contrôles [47].
La capacité de résister à l’envie de manger exige notamment un bon fonctionnement des réseaux neuronaux impliqués dans le contrôle de la prise alimentaire. Ces réseaux ont pour effet l’inhibition de réponses conditionnées suivant la présentation d’un signal annonçant une récompense alimentaire. Il s’agit là typiquement d’un choix intertemporel exigeant de prendre en compte à la fois le plaisir immédiat et les conséquences pour la santé à long terme. Une étude IRMf a circonscrit les aires cérébrales impliquées dans ce type de choix [48], en indiquant que les activations du cortex préfrontal diffèrent lorsque les individus choisissent des aliments sains et des « junk food ». Si la valorisation de l’aliment semble dans tous les cas faire intervenir le cortex préfrontal ventro-médian, le choix de l’aliment sain fait aussi intervenir le cortex préfrontal dorso-latéral, lequel impliquerait la modulation de la valeur du signal.
Nous avons indiqué précédemment que la valorisation excessive de la « junk food » peut conduire à une consommation irrésistible et prédisposer également à une plus grande sensibilité aux publicités alimentaires. D’autres travaux utilisant l’IRMf et la tomographie par émission de positons (TEP) ont en effet permis de mettre en évidence des dysfonctionnements dans les réseaux neuronaux impliqués dans le conditionnement et le contrôle cognitif [49, 50]. Ces études ont indiqué une corrélation négative entre l’indice de masse corporelle et l’activité du cortex orbito-frontal et préfrontal dorso-latéral, ainsi qu’avec le volume de matière grise dans ces régions. Il a été signalé précédemment que le cortex préfrontal dorso-latéral intervenait dans la valorisation des options décalées dans le temps [40]. Le choix de l’aliment sain demanderait que l’individu exerce un contrôle sur soi pour inhiber la valorisation d’une récompense immédiate (liée à l’ingestion) afin de prendre en compte la récompense d’une alimentation saine sur le long terme. Loin d’une théorie de la rationalité substantive, l’IRMf permet d’informer de certaines spécificités de l’activité cérébrale des personnes souffrant d’obésité, suggérant une valorisation excessive du plaisir attendu de la consommation et un manque de contrôle lié à une faible valorisation des options futures. Ces résultats du point de vue de la prévention en santé publique suggérent une faible efficacité des messages de prévention de l’obésité basés sur la seule idée de bénéfices à long terme d’une alimentation saine.
Mise en perspective
D’autres études utilisant l’IRMf indiquent qu’un argumentaire valorisant les qualités gustatives des aliments généralement catégorisés comme sains pourrait s’avérer efficace [51, 52]. Pour l’ingestion d’un même produit alimentaire, les aires du cerveau associées (notamment) au plaisir de manger - plus particulièrement le striatum - présentent une activité significativement plus élevée lorsque des messages décrivant l’aliment comme délicieux sont présentés que lorsque les descriptifs sont neutres indiquant juste la composition de l’aliment [51]. Comme suggéré dans les travaux du Centre d’analyse stratégique [53], qui pour la première fois propose d’utiliser conjointement les sciences comportementales et du cerveau dans le cadre de la prévention en santé publique (notamment dans la lutte contre l’obésité), un tel résultat peut sembler une évidence. Toutefois, cette évidence n’est pour l’instant pas mise en œuvre : les messages sanitaires présentent en effet plus souvent des données factuelles à visée utilitariste et sanitaire sur les aliments que des incitations hédoniques ! Ce rapport propose donc d’envisager, pour les aliments gras et sucrés, des descriptions publicitaires neutres dont l’efficacité pourrait être mesurée grâce à l’IRMf (sans pour autant négliger les instruments plus classiques d’évaluation de l’efficacité des campagnes des prévention) [53].
Une autre expérience montre comment des aliments bien présentés (travail de mise en forme dans l’assiette par exemple) par rapport à des présentations d’aliments sans artifice semblent contribuer à la préparation de l’action motrice et à la sensation de plaisir avant la consommation (amygdale, striatum ventral, cortex cingulaire antérieur, cortex prémoteur). Cette activité est d’autant plus importante que les sujets, suite à un questionnaire préalable, sont classés comme sensibles à la récompense alimentaire, sensibilité qui est positivement corrélée à l’obésité [46]. Ces résultats suggèrent que les personnes souffrant d’obésité seraient particulièrement sensibles aux campagnes publicitaires valorisant des produits fortement caloriques, et ce d’autant que des efforts sont mis en œuvre pour les valoriser dans leur présentation. Là encore, de tels résultats, s’il peuvent apparaître comme une évidence, soutiennent des actions futures en faveur de présentations plus « neutres » des aliments gras et sucrés. De telles expériences suggèrent que les messages de prévention gagneraient à présenter des images appétissantes de fruits et légumes et à vanter leurs qualités hédoniques. Ainsi, à moyen terme, l’imagerie cérébrale fonctionnelle pourrait devenir une méthodologie complémentaire à celle utilisée dans l’élaboration et l’évaluation des campagnes de prévention en santé publique [53]. Toutefois, les résultats de neuro-imagerie ne seront utiles qu’à l’aune de travaux en psychologie sociale et différentielle - disciplines encore trop timidement convoquées en santé publique pour la première et quasiment absente pour la seconde - et de modèles théoriques permettant de lier les différents niveaux d’analyse du comportement [54].
Conclusion
L’observation des comportements humains dans leur quotidien « réel » et non supposé, ainsi que de récents travaux complémentaires utilisant l’imagerie cérébrale - et notamment l’IRMf - remettent en question le postulat de la rationalité substantive qui prévaut depuis des décennies en sciences économiques. Ces travaux apportent de nouveaux éléments de compréhension des mécanismes en jeu lors du traitement des informations dans le contexte de la prise de décision et montrent l’interdépendance émotion-rationalité dans le traitement cérébral [55].
L’utilisation de la neuro-imagerie contribue aussi à mieux comprendre ce qui amène certaines personnes à choisir la pomme et d’autres la barre chocolatée dans la situation que nous présentions en tout début d’article. Elle met en évidence des différences interindividuelles dans le traitement des informations qui dépassent la simple expression des préférences. S’il est clair que toutes les informations n’ont pas la même pertinence en fonction des individus, l’IRMf indique que celle-ci peut être en partie déterminée par leur facilité de traitement [37], les capacités de self-control [48] et de projection dans le temps [40] évidemment susceptibles de différer entre les individus. Plus spécifiquement, certaines populations, comme celle des personnes souffrant d’obésité, ne pourraient traiter les informations de manière similaire du fait de contraintes cérébrales fonctionnelles voire structurelles [45, 47, 49, 50]. Dans le cadre de la prévention de l’obésité ce constat demande de conduire des campagnes de prévention plus adaptées aux populations ciblées, notamment en utilisant les dernières avancées en psychologie différentielle et en imagerie cérébrale, non pour remplacer les travaux et méthodes qui sont employés, mais pour les compléter voire les préciser.
Conflit d’intérêts
Les auteurs déclarent n’avoir aucun conflit d’intérêts concernant les données publiées dans cet article.
Remerciements
La rédaction du présent article a bénéficié du soutien de la Fondation sport santé et développement durable.
à supposer qu’un choix puisse être « arrêté », tant une fois la décision prise, nous avons tendance à y repenser, la regrettant souvent quand celle-ci n’est pas la meilleure pour notre santé et que nous le savons.
Champ d’investigation qui explore les liens entre les activités des systèmes nerveux et endocrinien et les décisions économiques et financières.
Notons toutefois que la variabilité due à la technique d’imagerie cérébrale, aux caractéristiques de l’instrument de mesure et au traitement des données ne sont pas les seules à entrer en compte dans ce cas là. Celles qui sont liées aux différences (neuro)anatomiques entre individus ou encore à la répétition de la tâche et donc à la mise en place potentielle de processus d’apprentissage ne peuvent être éludées.
Références
- Laan LN van der, Ridder DTD de, Viergever MA, et al. The first taste is always with the eyes: A meta-analysis on the neural correlates of processing visual food cues. NeuroImage 2011 ; 55 : 55296–55303. [Google Scholar]
- Simmons WK, Martin A, Barsalou LW. Pictures of appetizing foods activate gustatory cortices for taste and reward. Cereb Cortex 2005 ; 15 : 1602–1608. [Google Scholar]
- Henrich J, Heine SJ, Norenzayan A. The weirdest people in the world? Behav Brain Sci 2010 ; 33 : 61–135. [Google Scholar]
- Bennett CM, Miller MB. How reliable are the results from functional magnetic resonance imaging? Ann NY Acad Sci 2010 ; 1191 : 133–155. [CrossRef] [Google Scholar]
- Von Neumann J, Morgenstern O. Theory of game and economic behavior. Princeton NJ : Princeton University Press, 1944. [Google Scholar]
- Savage LG. The foundations of statistics. New York : Dover Publications, 1954. [Google Scholar]
- Luce RD, Raiffa H. Games and decisions. New York : Wiley and Sons, 1957. [Google Scholar]
- Samuelson PA. A note on the pure theory of consumer’s behaviour. Economica 1938 ; 5 : 61–71. [CrossRef] [Google Scholar]
- Oullier O, Basso F. Embodied economics: how bodily information shapes the social coordination dynamics of decision-making. Philos T Roy Soc B 2010 ; 365 : 291–310. [CrossRef] [Google Scholar]
- Joule RV, Beauvois JL. La soumission librement consentie. Paris : PUF, 2007. [Google Scholar]
- Huguet P, Dumas F, Monteil JM. Competing for a desired reward in the stroop task : when attentional control is unconscious but effective versus conscious but ineffective. Can J Exp Psychol 2004 ; 58 : 153–167. [CrossRef] [PubMed] [Google Scholar]
- Simon HA. A behavioral model of rational choice. Q J Eco 1955 ; 69 : 99–118. [Google Scholar]
- Park CW. A seven-point scale and a decision maker’s simplifying strategy: an operationalized satisficing-plus model. Organ Behav Hum Perf 1978 ; 21 : 252–271. [CrossRef] [Google Scholar]
- Beach LR, Mitchell TR. A contingency model for the selection of decision strategies. Acad Manag Rev 1978 ; 3 : 439–449. [CrossRef] [Google Scholar]
- Christensen-Szalanski JJ. A further examination of the selection of problem-solving strategies: the effects of deadlines and analytic aptitudes. Organ Behav Hum Perf 1980 ; 25 : 107–122. [CrossRef] [Google Scholar]
- Ellsberg D. Risk, ambiguity, and the Savage axioms. Quart J Econ 1961 ; 75 : 643–669. [Google Scholar]
- Tversky A, Kahneman D. Judgment under uncertainty: Heuristics and biases. Science 1974 ; 185 : 1124–1131. [CrossRef] [PubMed] [Google Scholar]
- Kahneman D, Tversky A. Prospect theory: An analysis of decision under risk. Econometrica 1979 ; 47 : 263–291. [CrossRef] [Google Scholar]
- Camerer C, Loewenstein G, Prelec D. Neuroeconomics: How neuroscience can inform economics. J Econ Lit 2005 ; 43 : 9–64. [CrossRef] [Google Scholar]
- Sanfey AG, Loewenstein G, McClure SM, et al. Neuroeconomics: cross-currents in research on decision-making. Trends Cogn Sci 2006 ; 10 : 108–116. [CrossRef] [PubMed] [Google Scholar]
- Glimcher PW. Foundations of neuroeconomic analysis. Oxford : Oxford University Press, 2010. [CrossRef] [Google Scholar]
- Oullier O, Kelso JAS. Neuroeconomics and the metastable brain. Trends Cogn Sci 2006 ; 10 : 353–354. [CrossRef] [PubMed] [Google Scholar]
- Haber SN, Knutson B. The reward circuit: linking primate anatomy and human imaging. Neuropsychopharmacol 2010 ; 35 : 4–26. [CrossRef] [Google Scholar]
- Wang GJ, Volkow ND, Telang F, et al. Exposure to appetitive food stimuli markedly activates the human brain. Neuroimage 2004 ; 21 : 1790–1797. [CrossRef] [PubMed] [Google Scholar]
- Hervé D, Matamales M, Stipanovich A, et al. Un nouveau mécanisme par lequel la récompense et les drogues modifient la chromatine dans les neurones. Med Sci (Paris) 2008 ; 24 : 1027–1029. [CrossRef] [EDP Sciences] [PubMed] [Google Scholar]
- Knutson B, Peterson R. Neurally reconstructing expected utility. Game Econ Behav 2005 ; 52 : 305–315. [Google Scholar]
- Schultz W, Dayan P, Montague PR. A neural substrate of prediction and reward. Science 1997 ; 275 : 1593–1599. [CrossRef] [PubMed] [Google Scholar]
- Camerer CF. Wanting, liking, and learning: neuroscience and paternalism. U Chicago Law Rev 2006 ; 73 : 87–110. [Google Scholar]
- Berridge KC, Robinson TE. Parsing reward. Trends Neurosci 2003 ; 26 : 507–513. [CrossRef] [PubMed] [Google Scholar]
- Berridge KC. The debate over dopamine’s role in reward: the case for incentive salience. Psychopharmacology 2007 ; 191 : 391–431. [CrossRef] [PubMed] [Google Scholar]
- Berridge KC, Robinson TE. What is the role of dopamine in reward: hedonic impact, reward learning, or incentive salience? Brain Res Rev 1998 ; 28 : 309–369. [Google Scholar]
- Brauer LH, Cramblett MJ, Paxton DA, et al. Haloperidol reduces smoking of both nicotine-containing and denicotinized cigarettes. Psychopharmacologia 2001 ; 159 : 31–37. [CrossRef] [Google Scholar]
- Evans AH, Pavese N, Lawrence AD, et al. Compulsive drug use linked to sensitized ventral striatal dopamine transmission. Ann Neurol 2006 ; 59 : 852–858. [CrossRef] [PubMed] [Google Scholar]
- Voon V, Pessiglione M, Brezing C, et al. Mechanisms underlying dopamine-mediated reward bias in compulsive behaviors. Neuron 2010 ; 65 : 135–142. [CrossRef] [PubMed] [Google Scholar]
- Finlayson G, King N, Blundell J. The role of implicit wanting in relation to explicit liking and wanting for food: Implications for appetite control. Appetite 2008 ; 50 : 120–127. [CrossRef] [PubMed] [Google Scholar]
- Kahneman D, Tversky A. Choices, values, and frames. Am Psychol 1984 ; 39 : 341–350. [Google Scholar]
- Smith K, Dickhaut J, McCabe K, et al. Neuronal substrates for choice under ambiguity, risk, gains, and losses. Manage Sci 2002 ; 48 : 711–718. [CrossRef] [Google Scholar]
- Damasio AR. Descartes’ Error. New York : Putnam. Trad. fr. de M. Blanc. L’erreur de Descartes. La raison des émotions. Paris : Odile Jacob, 1994. [Google Scholar]
- Laibson DI, Repetto A, Tobacman J. Self control and retirement savings. Brook Papers Econ Act 1998 ; 1 : 91–172. [CrossRef] [Google Scholar]
- McClure SM, Laibson DI, Loewenstein G, et al. Separate neural systems value immediate and delayed monetary rewards. Science 2004 ; 306 : 503–507. [CrossRef] [PubMed] [Google Scholar]
- Dehaene S, Dehaene-Lambertz G, Cohen L. Abstract representations of numbers in the animal and human brain. Trends Neurosci 1998 ; 21 : 355–361. [CrossRef] [PubMed] [Google Scholar]
- Plassmann H, O’Doherty J, Rangel A. Orbitofrontal cortex encodes willingness to pay in everyday economic transactions. J Neurosci 2007 ; 27 : 9984–9988. [CrossRef] [PubMed] [Google Scholar]
- Plassmann H, O’Doherty JP, Rangel A. Appetitive and aversive goal values are encoded in the medial orbitofrontal cortex at the time of decision making. J Neurosci 2010 ; 30 : 10799–10808. [CrossRef] [PubMed] [Google Scholar]
- Berridge KC. ‘Liking’ and ‘wanting’ food rewards: brain substrates and roles in eating disorders. Physiol Behav 2009 ; 97 : 537–550. [Google Scholar]
- Stice E, Spoor S, Bohon C, et al. Relation of reward from food intake and anticipated food intake to obesity: a functional magnetic resonance imaging study. J Abnorm Psychol 2008 ; 117 : 924–935. [CrossRef] [PubMed] [Google Scholar]
- Davis C, Strachan S, Berkson M. Sensitivity to reward: implications for overeating and overweight. Appetite 2004 ; 42 : 131–138. [CrossRef] [PubMed] [Google Scholar]
- Weller RE, Cook III EW,, Avsar KB, et al. Obese women show greater delay discounting than healthy-weight women. Appetite 2008 ; 51 : 563–569. [CrossRef] [PubMed] [Google Scholar]
- Hare TA, Camerer CF, Rangel A. Self-control in decision-making involves modulation of the vmPFC valuation system. Science 2009 ; 324 : 646–648. [CrossRef] [PubMed] [Google Scholar]
- Volkow ND, Wang GJ, Telang F, et al. Low dopamine striatal D2 receptors are associated with prefrontal metabolism in obese subjects: possible contributing factors. Neuroimage 2008 ; 42 : 1537–1543. [CrossRef] [PubMed] [Google Scholar]
- Walther K, Birdsill AC, Glisky EL, et al. Structural brain differences and cognitive functioning related to body mass index in older females. Hum Brain Mapp 2010 ; 31 : 1052–1064. [CrossRef] [PubMed] [Google Scholar]
- Grabenhorst F, Rolls ET, Bilderbeck A. How cognition modulates affective responses to taste and flavor: top-down influences on the orbitofrontal and pregenual cingulate cortices. Cereb Cortex 2008 ; 18 : 1549–1559. [CrossRef] [PubMed] [Google Scholar]
- Passamonti L, Rowe JB, Schwarzbauer C, et al. Personality predicts the brain’s response to viewing appetizing foods: the neural basis of a risk factor for overeating. J Neurosci 2009 ; 29 : 43–51. [CrossRef] [PubMed] [Google Scholar]
- Oullier O, Sauneron S. Nouvelles approches de la prévention en santé publique : l’apport des sciences comportementales, cognitives et des neurosciences (Rapport n°25 du Centre d’analyse stratégique). Paris : La Documentation Française, 2010 : 192 p. [Google Scholar]
- Oullier O, Kirman AP, Kelso JAS. The coordination dynamics of economic decision making: a multilevel approach to social neuroeconomics. IEEE Trans Neural Syst Rehab Eng 2008 ; 16 : 557–571. [CrossRef] [Google Scholar]
- Oullier O. The useful brain : Why neuroeconomics might change our views on rationality and a couple of other things. In: Michel-Kerjan E, Slovic P, eds. The irrational economist : making decisions in a dangerous world. New York : Public Affairs, 88–96. [Google Scholar]
Liste des figures
|
Figure 1. Méta-analyse d’études sur la vision de nourriture utilisant l’imagerie par résonance magnétique fonctionnelle. A. Activité significativement plus élevée lorsque l’individu voit de la nourriture en comparaison d’objets non alimentaires (rouge). B. Activité significativement plus élevée lorsque l’individu voit de la nourriture en état de faim versus en état de satiété (bleu), et activité significativement plus élevée lorsque la nourriture vue est plus calorique que « diététique » (moins calorique ; vert) (adapté de [1]). |
| Dans le texte | |
|
Figure 2. Schéma des principales structures et connexions composant le circuit de la récompense. Flèches rouges : efférences du cortex préfrontal ventromédian (vmPFC) vers le striatum ventral ; flèches oranges : efférences de la partie dorsale du cortex cingulaire antérieur (dACC) vers le striatum ventral ; flèches jaunes : efférences du cortex préfrontal dorsal (dPFC) vers le striatum ventral ; flèches marrons : principales connexions du circuit de la récompense (adapté de [23]). |
| Dans le texte | |
|
Figure 3. Risque et incertitude. La tâche du participant est de choisir entre deux paris en fonction des distributions de probabilités et de revenus pour chacune des billes constituant l’urne. Les chiffres au-dessus des flèches indiquent la distribution des billes rouges, noires et jaunes dans les urnes. Les chiffres en dessous des flèches indiquent les gains en dollars. Deux conditions sont présentées : A. Les sujets doivent choisir entre deux paris risqués sous forme de gain. Le pari sur la gauche a une variance plus faible et est considéré comme moins risqué. B. Les sujets doivent choisir entre un pari présentant une part d’incertitude (à gauche) et le même pari risqué que la condition précédente en A (à droite). Les flèches obliques indiquent qu’il y a un total de 60 billes noires et jaunes dans le pari 1. Le nombre précis de billes noires et vertes n’est pas connu (adapté de [37]). |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.