")
")
| Issue |
Med Sci (Paris)
Volume 25, Number 12, Décembre 2009
Anticorps monoclonaux en thérapeutique
|
|
|---|---|---|
| Page(s) | 1033 - 1038 | |
| Section | I - De la conception à la production | |
| DOI | https://doi.org/10.1051/medsci/200925121033 | |
| Published online | 15 décembre 2009 | |
Protéine de fusion ou anticorps monoclonal
Quel biomédicament choisir dans une maladie inflammatoire ?
Recombinant proteins or monoclonal antibodies: comparative properties and interest in rheumatoid arthritis
Centre national de reference « Maladies auto-immunes systémiques rares », service de rhumatologie, CHU de Strasbourg, Hôpital de Hautepierre, 1, avenue Molière, BP 83049, 67098 Strasbourg Cedex, France
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Résumé
Les options thérapeutiques dans les maladies inflammatoires ont été bouleversées par l’introduction de molécules biologiques ciblant les cytokines et/ou la réponse immune impliquées dans la pathologie. Ces agents sont soit des anticorps monoclonaux contre une « cible » thérapeutique (cytokine ou cellule), soit des protéines recombinantes reconnaissant aussi ces cibles. Cette question est particulièrement bien illustrée dans les maladies inflammatoires comme la polyarthrite rhumatoïde (PR) ou des maladies auto-immunes, où plusieurs anticorps monoclonaux et protéines recombinantes humaines chimériques (étanercept ciblant le TNF dans la PR, ou abatacept ciblant CTLA-4 et inactivant les lymphocytes T) ont été développés et utilisés avec succès. Dans le cas de la PR que nous discuterons particulièrement, cinq molécules sont disponibles et leurs propriétés et efficacité ont donc été comparées. Nous discuterons les paramètres pris en compte dans le choix de l’une ou l’autre de ces molécules, qu’ils soient liés à la structure de la molécule (relations structure-fonction, pharmacocinétique, affinité pour sa cible, mécanismes d’action) ou à la susceptibilité propre au patient ou à la maladie. Une connaissance approfondie de ces caractéristiques et les progrès de la bio-ingénierie permettront d’optimiser leur utilisation mais aussi d’imaginer les molécules du futur.
Abstract
Therapeutics options for inflammatory diseases, such as rheumatoid arthritis (RA), have increased tremendously in the past decade with the introduction of biologic therapies, such as monoclonal antibodies or recombinant fusion proteins. These have proven to be highly successful in treating inflammatory or autoimmune diseases, by blocking certain key molecules involved in the pathogenesis of the illness, cytokines (TNF) or immune coactivators (CTLA-4). Thus in rheumatoid arthritis, TNF can be neutralized both by monoclonal antibodies (adalimumab, infliximab) or recombinant inhibitors such as etanercept or CTLA-4 Ig abatacept. All have been marketed and proven to be highly effective in the treatment of RA, and we will discuss parameters which are taken into account to select monoclonal antibody or recombinant inhibitors. These include drug-related (target affinity, pharmacokinetics, mechanisms of action, etc.) and patient - (efficacy and side effects) or disease-related characteristics. Their impact on current clinical practice and future trends are discussed.
© 2009 médecine/sciences - Inserm / SRMS
Pour agir de façon ciblée dans une maladie inflammatoire, le plus simple est souvent de développer un anticorps monoclonal (Acm) contre une « cible » thérapeutique qui peut être une cytokine ou une cellule intervenant de façon importante dans la réponse immunitaire [1]. Cependant, le choix d’une protéine de fusion est une option parfois pertinente comme l’illustre l’exemple des traitements de la polyarthrite rhumatoïde (PR) avec la mise sur le marché de l’étanercept (Enbrel®), une molécule anti-TNF (tumor necrosis factor) et d’un inhibiteur de la costimulation (abatacept : Orencia®).
Le choix d’une protéine de fusion pose de manière schématique deux questions : (1) comment choisir la cible thérapeutique de cette molécule ? (2) Pourquoi choisir plutôt une protéine de fusion qu’un Acm ?
Comment choisir la cible thérapeutique d’une protéine de fusion ?
Le principe général est le plus souvent d’essayer de mimer un mécanisme de régulation physiologique, ce qui est le cas pour l’étanercept et l’abatacept dans la polyarthrite rhumatoïde (PR).
Étanercept
L’étanercept est une protéine recombinante humaine dimérique composée de la portion extracellulaire du récepteur de type 2 (TNFR2/p75) du TNF fusionnée à un fragment Fc (avec une région charnière incomplète sans CH2) d’une IgG1 humaine qui comprend 13 résidus O-glycosyl et 11 résidus proline. Ainsi, l’étanercept peut mimer partiellement l’effet du récepteur TNFR2 (p75) soluble qui est un antagoniste naturel du TNF. En fait, rien ne permet réellement d’argumenter le choix du récepteur de type 2 (TNFR2/p75) par rapport au récepteur de type 1 (TNFR1/p55) car ces deux récepteurs solubles (issus du clivage des récepteurs membranaires) ont globalement les mêmes capacités d’inhibition du TNF soluble et membranaire, même s’il s’agit d’un point qui a été peu étudié. Les différentes protéines de fusion TNFR-IgG utilisées en thérapeutique ont des affinités pour le TNF supérieures à celles des formes natives solubles de TNFR [2].
Avant l’étanercept, une autre protéine de fusion, le lénercept (qui est un dimère formé de la portion extracellulaire du TNFR1/p55 et d’un domaine Fc d’IgG1 avec une région charnière complète) a été évaluée dans le choc septique, la sclérose en plaques et la polyarthrite rhumatoïde sans montrer d’efficacité significative [3–6]. L’échec de cette molécule pourrait être lié à sa glycosylation qui peut modifier sa pharmacocinétique et sa bioactivité [7] ou à son pouvoir immunogène responsable de l’apparition d’anticorps anti-lénercept [6]. Peu d’études comparatives ont été menées mais il a été démontré que le TNFR soluble p75-Fc fixe et relargue cent fois plus vite le TNF soluble que ne le fait le TNFR soluble p55-Fc [8]. Dans un modèle d’arthrite lié à la surexpression du TNF membranaire murin chez des souris transgéniques, deux récepteurs solubles (p55-PEG et p75-Fc) semblent avoir une efficacité comparable malgré des différences pharmacocinétiques [9].
Abatacept
L’abatacept est une protéine recombinante humaine composée d’un dimère des domaines extracellulaires du CTLA-41 (cytotoxic T-lymphocyte antigen 4, ou CD152) humain (relié par des ponts disulfures) fusionnés aux fragments Fc modifiés d’une IgG1 humaine comportant la région charnière et les domaines CH2-CH3. Les modifications de la région charnière du fragment Fc sont des mutations remplaçant les cystéines en position 130, 136 et 139 par des sérines et la proline en position 148 par une sérine, ce qui permet la fixation aux récepteurs des fragments Fc (FcγR) sans que cette molécule n’induise de cytotoxicité par ADCC (antibody-dependent cellular cytotoxycity) ou CDC (complement-dependent cytotoxycity) [10] (→).
(→) Voir R. Abès et al., page 1011
L’abatacept permet d’inactiver les lymphocytes T car il entre en compétition avec CD28 pour la liaison aux molécules B7 (B7.1 et.2) exprimées par les cellules présentatrices de l’antigène. Physiologiquement, ces cellules, via la fixation de CD28 sur B7, envoient un premier signal de costimulation activateur aux lymphocytes T, ce qui induit l’expression de CTLA-4 qui vient interrompre cette activation en délivrant un signal inhibiteur par sa fixation sur B7. La fixation de CTLA-4 est facilitée par son affinité pour B7 qui est 500 à 2 500 fois supérieure à celle de CD28. En pratique, l’abatacept permet de reproduire ce phénomène inhibiteur physiologique, ce qui est mis à profit dans des maladies auto-immunes caractérisées par une activation lymphocytaire excessive. Cependant, l’abatacept, efficace dans la PR, n’avait pas l’efficacité espérée dans la prévention du rejet de greffe. En fait, l’abatacept ne bloque pas suffisamment la costimulation B7-CD28 car il a une affinité trop faible pour B7.2 (CD86).
En réponse à ce problème, une molécule analogue à l’abatacept a été créée, le bélatacept (LEA 29Y), qui a deux mutations remplaçantes situées dans les boucles CDR1 (complementary determining regions) de la portion extracellulaire de CTLA-4, ce qui lui confère une avidité quatre fois plus importante pour B7.2 (CD86) et deux fois plus forte pour B7.1 (CD80) [11]. Ainsi, l’exemple de ces deux molécules, l’abatacept et le bélatacept, démontre qu’une stratégie fondée sur les modifications de structure permet d’obtenir la molécule la plus efficace possible dans des situations bien définies avec, si possible, une meilleure tolérance [12].
Pourquoi une molécule de fusion plutôt qu’un Acm ?
Cette question peut être illustrée par les anti-TNF qui regroupent actuellement quatre Acm (infliximab, adalimumab, certolizumab, golimumab) et une protéine de fusion (étanercept) (Figure 1). En pratique, le rapport bénéfice/risque de ces cinq molécules est assez comparable mais il y a quelques différences notables (Tableau I).
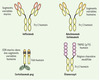 |
Figure 1. Schéma des cinq molécules anti-TNF utilisées en pathologie humaine. PEG : polyéthylène glycol ; CDR : complémentation determining region. |
Caractéristiques des différents anti-TNF. PEG : polyéthylène glycol. TNF sol : tumor necrosis factor soluble ; TNF mb : tumor necrosis factor membranaire ; CDC : complement-dependent cytotoxicity ; ADCC : antibody-dependent cellular cytotoxicity ; LT : lymphotoxine.
Efficacité thérapeutique
L’efficacité de ces molécules (infliximab, adalimumab) est équivalente dans la PR et les spondylarthropathies mais l’étanercept est moins efficace que les Acm dans les affections granulomateuses (maladie de Crohn, granulomatose de Wegener, sarcoïdose, etc.) et à un degré moindre dans le psoriasis cutané et certaines uvéites sévères [13].
Différences de tolérance traduisant des caractéristiques différentes
La tolérance est assez proche mais il y a des différences intéressantes avec moins d’infections tuberculeuses et probablement moins de phénomènes d’auto-immunisation (induction d’auto-anticorps et de lupus) sous étanercept [14, 15] (→). Ces différences traduisent certainement des caractéristiques (structure et affinité/avidité), des modes d’administration et des mécanismes d’action différents (Tableau I). L’analyse de ces différences est complexe car elles sont en théorie très nombreuses, et les études qui s’y sont intéressées sont peu nombreuses et hétérogènes. Néanmoins, parmi ces différences, quelques-unes méritent d’être soulignées.
(→) voir M. Pallardy, page 1130 ; E. Rigal et al., page 1135
• Affinité de la molécule de fusion ou de l’Acm pour les différentes formes de TNF
Les Acm fixent le TNF soluble monomérique ou trimérique ainsi que le TNF membranaire. L’étanercept fixe avec la même affinité le TNF soluble (mais uniquement dans sa forme trimérique) mais pourrait avoir une affinité moins forte pour le TNF membranaire qu’il reconnaît par des épitopes différents de ceux reconnus par les Acm [16, 17]. En fait, des travaux récents semblent montrer que l’infliximab, l’adalimumab, le certolizumab et l’étanercept peuvent fixer le TNF membranaire avec une affinité comparable mais cette affinité est globalement moins forte que pour le TNF soluble. L’un des points les plus importants pour comprendre la neutralisation du TNF est la caractérisation des complexes TNF/anti-TNF dont la structure peut être variable. D’une part, les Acm bivalents fixent deux trimères de TNF soluble alors que l’étanercept n’en fixe qu’un, mais l’avidité de ces molécules est comparable voire supérieure pour l’étanercept [18]. D’autre part, trois molécules d’infliximab ou d’adalimumab peuvent se fixer sur une molécule de TNF membranaire alors que l’étanercept n’en fixe qu’une ; mais surtout l’étanercept, qui est monovalent, ne relie pas (cross link) les TNF membranaires. De plus, les Acm anti-TNF peuvent reconnaître les complexes TNF/TNFR, ce que ne fait pas l’étanercept [19, 20]. Ces différences stœchiométriques peuvent avoir des conséquences fonctionnelles variées.
Alors que les Acm empêchent l’accès aux deux récepteurs du TNF (TNFR1 et R2) en bloquant les deux formes (soluble et membranaire) de TNF, l’étanercept peut préserver partiellement l’activation du TNFR2 qui se fait préférentiellement via la fixation du TNF membranaire. Ce TNF membranaire a une affinité supérieure pour le TNFR2 avec lequel il forme un complexe moins stable que le complexe TNF soluble-TNFR1 p55 [21]. Cette différence pourrait expliquer la moindre efficacité de l’étanercept dans les granulomatoses mais aussi le risque réduit de tuberculose [14, 15, 22]. En effet, il a été démontré que le TNFR2 et le TNF membranaire jouent un rôle important dans la formation du granulome et dans la défense antituberculeuse [23, 24]. Cette interaction différente avec le TNF membranaire peut avoir d’autres conséquences, notamment pour la défense antitumorale. Il a été démontré, dans des modèles murins, que le TNFR2, exprimé par les macrophages et activé par du TNF membranaire, pouvait avoir une puissante action antitumorale car il permet la libération de NO (nitric oxid) macrophagique qui va agir sur les cellules endothéliales, et cela indépendamment des lymphocytes B et T [25].
La fixation sur le TNF membranaire peut provoquer un phénomène appelé reverse signaling parce que le signal est déclenché dans la cellule exprimant la cytokine [26, 27]. Cela peut induire l’apoptose et l’inhibition de la production de cytokines mais aussi l’activation de cette cellule et la surexpression de sélectines [17, 28]. Cette fixation peut aussi moduler les fonctions des lymphoyctes T CD4 et CD8 [29]. L’étanercept n’induit pas (ou peu) l’apoptose des cellules exprimant du TNF membranaire car cet anti-TNF se fixe sans établir de lien (cross link) entre les molécules de TNF [30–33]. Cependant, dans certaines conditions, l’étanercept est bien capable d’induire une apoptose, en particulier celle des monocytes et des macrophages synoviaux dans la PR mais aussi des cellules dendritiques dans le psoriasis cutané [34–37]. En réalité, chez l’homme, les effets pro-apoptotiques des anti-TNF sont difficiles à analyser et à interpréter. Les différences observées pourraient être la conséquence in vitro d’une variabilité liée aux cellules ou aux tissus et aux techniques utilisées (concentration d’anti-TNF, durée d’incubation, conditions de culture, costimulation, analyse de l’apoptose, etc.) [38].
• Fixation des lymphotoxines
Seul l’étanercept peut fixer les lymphotoxines LTα3 soluble et LTα2β1 membranaire sans que l’on sache avec quelle affinité. Les conséquences cliniques de cette interaction sont mal connues chez l’homme mais les lymphotoxines jouent un rôle particulier dans le développement du système lymphoïde et dans le fonctionnement du système immunitaire, avec probablement un effet sur la maturation des lymphocytes B [39, 40].
• Élimination des molécules anti-TNF
Elle peut varier selon la molécule qui favorise des complexes TNF/anti-TNF de taille et de stabilité variables. De plus, l’étanercept est capable de relarguer le TNF soluble après fixation [16]. En effet, des complexes TNF soluble/étanercept, qui persistent longtemps chez l’homme, ont la possibilité de relarguer du TNF bioactif, ce qui en fait de potentiels « porteurs de TNF » avec d’éventuelles conséquences clinico-biologiques [28, 41–43]. Néanmoins, ce phénomène de relargage du TNF est difficile à expliquer compte tenu de l’affinité/avidité de l’étanercept qui est au moins comparable à celle des anticorps monoclonaux. L’élimination de l’étanercept dépend aussi d’autres facteurs, en particulier de sa capacité à se fixer au récepteur néonatal des fragments Fc (FcRn) (→). La demi-vie plasmatique plus prolongée des Acm anti-TNF (infliximab, adalimumab) s’explique notamment par le fait que ces molécules fixent le FcRn alors que l’étanercept ne le fixe pas.
(→) voir C. Magdelaine-Beuzelin et al., page 1053
• Cytotoxicité due au fragment Fc
En théorie, les différentes molécules anti-TNF (sauf le certolizumab qui est un Fab monovalent) peuvent induire un effet de cytotoxicité lié à leurs fragments Fc. Une CDC, et surtout une ADCC, peuvent être observées avec l’infliximab et l’adalimumab et à un degré moindre avec l’étanercept [26, 35, 44]. Cependant, dans une étude in vitro utilisant trois anti-TNF à des concentrations cliniques, aucune cytotoxicité n’a été détectée sur des cellules (sang total) stimulées par Mycobacterium tuberculosis [45, 46]. Dans un autre travail récent, aucun TNF n’est capable d’induire la CDC de monocytes stimulés in vitro par des molécules induisant l’expression du TNF membranaire [18]. En fait, aucune donnée in vivo chez les patients n’a réellement évalué l’impact de ces mécanismes de cytotoxicité dans les maladies inflammatoires.
• Effets immunomodulateurs des anti-TNF
Ils agissent en particulier sur la migration cellulaire mais ils peuvent avoir d’autres effets amplificateurs ou suppresseurs de la réponse immunitaire. L’effet le plus original est la restauration de la fonction des lymphocytes T régulateurs (CD4+CD25+Foxp3+) observée avec l’infliximab [47, 48]. Ces molécules ont aussi un impact sur les lymphocytes B car les anti-TNF peuvent réduire (inconstamment) la production d’auto-anticorps (facteurs rhumatoïdes et Ac antipeptides citrullinés) dans la PR alors que par d’autres mécanismes, ils favorisent l’apparition d’Ac antinucléaires et d’Ac antiADN natifs (surtout d’isotype IgM). L’étanercept, qui inhibe également les lymphotoxines, a certainement des effets particuliers sur les lymphocytes B mais qui sont à ce jour mal connus [39]. En revanche, l’effet inhibiteur de l’étanercept sur l’activation et la prolifération des lymphocytes T semble inférieur à celui des Acm, notamment sur la réponse antituberculeuse [44, 49].
Conclusion
Les protéines de fusion ont des originalités de structure et de fonction qui en font des outils thérapeutiques particulièrement performants dans les maladies inflammatoires comme la polyarthrite rhumatoïde, les arthrites juvéniles idiopathiques, les spondylarthropathies, le psoriasis et potentiellement d’autres maladies auto-immunes ou auto-inflammatoires. Une connaissance approfondie des relations entre leur structure et leur fonction peut permettre d’optimiser leur utilisation mais aussi d’imaginer les molécules du futur.
Conflit D’Intérêts
L’auteur déclare avoir participé à des interventions ponctuelles pour les entreprises Wyeth, Abbott, Schering Plough, Roche, Shugai, Bristol-Myers Squibb.
CTLA-4 fait partie de la superfamille des immunoglobiulines ; elle est induite lors de l’activation des lymphocytes T en réponse à la fixation de l’antigène ; homologue à la molécule d’activation lymphocytaire T CD28, elle a comme ligands B27-1 et B27-2, et cette liaison délivre un signal inhibiteur de l’activation lymphocytaire (arrêt de prolifération, inhibition de la synthèse d’IL-2).
Références
- Choy EH, Panayi GS. Cytokine pathways and joint inflammation in rheumatoid arthritis. N Engl J Med 2001; 344 : 907–16. [Google Scholar]
- Haak-Frendscho M, Marsters SA, Mordenti J, et al. Inhibition of TNF by a TNF receptor immunoadhesin. Comparison to an anti-TNF monoclonal antibody. J Immunol 1994; 152 : 1347–53. [Google Scholar]
- Butty VL, Roux-Lombard P, Garbino J, et al. Geneva Sepsis Network. Anti-inflammatory response after infusion of p55 soluble tumor necrosis factor receptor fusion protein for severe sepsis. Eur Cytokine Netw 2003; 14 : 15–9. [Google Scholar]
- Abraham E, Laterre PF, Garbino J, et al. Lenercept (p55 tumor necrosis factor receptor fusion protein) in severe sepsis and early septic shock: a randomized, double-blind, placebo-controlled, multicenter phase III trial with 1,342 patients. Crit Care Med 2001; 29 : 503–10. [Google Scholar]
- Rau R, Sander O, van Riel P, et al. Intravenous human recombinant tumor necrosis factor receptor p55-Fc IgG1 fusion protein Ro 45-2081 (lenercept): a double blind, placebo controlled dose-finding study in rheumatoid arthritis. J Rheumatol 2003; 30 : 680–90. [Google Scholar]
- TNF neutralization in MS: results of a randomized, placebo-controlled multicenter study. The Lenercept Multiple Sclerosis Study Group and The University of British Columbia MS/MRI Analysis Group. Neurology 1999; 53 : 457–65. [Google Scholar]
- Keck R, Nayak N, Lerner L, et al. Characterization of a complex glycoprotein whose variable metabolic clearance in humans is dependent on terminal N-acetylglucosamine content. Biologicals 2008; 36 : 49–60. [Google Scholar]
- Evans TJ, Moyes D, Carpenter A, et al. Protective effect of 55- but not 75-kD soluble tumor necrosis factor receptor-immunoglobulin G fusion proteins in an animal model of gram-negative sepsis. J Exp Med 1994; 180 : 2173–9. [Google Scholar]
- Edwards CK 3rd, Bendele AM, Reznikov LI, et al. Soluble human p55 and p75 tumor necrosis factor receptors reverse spontaneous arthritis in transgenic mice expressing transmembrane tumor necrosis factor alpha. Arthritis Rheum 2006; 54 : 2872–85. [Google Scholar]
- Davis PM, Abraham R, Xu L, et al. Abatacept binds to the Fc receptor CD64 but does not mediate complement-dependent cytotoxicity or antibody-dependent cellular cytotoxicity. J Rheumatol 2007; 34 : 2204–10. [Google Scholar]
- Vincenti F, Larsen C, Durrbach A, et al. Costimulation blockade with belatacept in renal transplantation. N Engl J Med 2005; 353 : 770–81. [Google Scholar]
- Larsen CP, Pearson TC, Adams AB, et al. Rational development of LEA29Y (belatacept), a high-affinity variant of CTLA4-Ig with potent immunosuppressive properties. Am J Transplant 2005; 5 : 443–53. [Google Scholar]
- Tracey D, Klareskog L, Sasso EH, et al. Tumor necrosis factor antagonist mechanisms of action: a comprehensive review. Pharmacol Ther 2008; 117 : 244–79. [Google Scholar]
- Furst DE, Wallis R, Broder M, Beenhouwer DO. Tumor necrosis factor antagonists: different kinetics and/or mechanisms of action may explain differences in the risk for developing granulomatous infection. Semin Arthritis Rheum 2006; 36 : 159–67. [Google Scholar]
- Tubach F, Salmon D, Ravaud P, et al. Risk of tuberculosis is higher with anti-tumor necrosis factor monoclonal antibody therapy than with soluble tumor necrosis factor receptor therapy: The three-year prospective french research axed on tolerance of biotherapies registry. Arthritis Rheum 2009; 60 : 1884–94. [Google Scholar]
- Scallon B, Cai A, Solowski N, et al. Binding and functional comparisons of two types of tumor necrosis factor antagonists. J Pharmacol Exp Ther 2002; 301 : 418–26. [Google Scholar]
- Nesbitt AM, Fossati G, Brown DT. Comparison of certolizumab pegol, etanercept, adalimumab and infliximab : effect on lipopolysaccharide-induced cytokine production by human peripheral blood monocytes. Am J Gastroenterol 2006; 101 : S420–70. [Google Scholar]
- Kaymakcalan Z, Sakorafas P, Bose S, et al. Comparisons of affinities, avidities, and complement activation of adalimumab, infliximab, and etanercept in binding to soluble and membrane tumor necrosis factor. Clin Immunol 2009; 131 : 308–16. [Google Scholar]
- Gramlick A, Fossati G, Nesbitt AM. Neutralization of soluble and membrane tumor necrosis factor-alpha (TNF-alpha) by infliximab, adalimumab, or certolizumab pegol using P55 or P75 TNF-alpha receptor specific bioassays. Gastroenterology 2006; 130 : A697. [Google Scholar]
- Kaymakcalan Z, Kalghatgi L, Xiong L. Differential TNF-neutralizing potencies of adalimumab, etanercept and infliximab. Ann Rheum Dis 2006; 65 : 458. [Google Scholar]
- Kassiotis G, Kollias G. Uncoupling the pro-inflammatory from the immunosuppressive properties of tumor necrosis factor (TNF) at the p55 TNF receptor level: implications for pathogenesis and therapy of autoimmune demyelination. J Exp Med 2001; 193 : 427–34. [Google Scholar]
- Wallis RS. Mathematical modeling of the cause of tuberculosis during tumor necrosis factor blockade. Arthritis Rheum 2008; 58 : 947–52. [Google Scholar]
- Olleros ML, Guler R, Corazza N, et al. Transmembrane TNF induces an efficient cell-mediated immunity and resistance to Mycobacterium bovis bacillus Calmette-Guérin infection in the absence of secreted TNF and lymphotoxin-alpha. J Immunol 2002; 168 : 3394–401. [Google Scholar]
- Saunders BM, Tran S, Ruuls S, et al. Transmembrane TNF is sufficient to initiate cell migration and granuloma formation and provide acute, but not long-term, control of Mycobacterium tuberculosis infection. J Immunol 2005; 174 : 4852–9. [Google Scholar]
- Zhao X, Mohaupt M, Jiang J, et al. Tumor necrosis factor receptor 2-mediated tumor suppression is nitric oxide dependent and involves angiostasis. Cancer Res 2007; 67 : 4443–50. [Google Scholar]
- Mitoma H, Horiuchi T, Hatta N, et al. Infliximab induces potent anti-inflammatory responses by outside-to-inside signals through transmembrane TNF-alpha. Gastroenterology 2005; 128 : 376–92. [Google Scholar]
- Kirchner S, Holler E, Haffner S, et al. Effect of different tumor necrosis factor (TNF) reactive agents on reverse signaling of membrane integrated TNF in monocytes. Cytokine 2004; 28 : 67–74. [Google Scholar]
- Suffredini AF, Reda D, Banks SM, et al. Effects of recombinant dimeric TNF receptor on human inflammatory responses following intravenous endotoxin administration. J Immunol 1995; 155 : 5038–45. [Google Scholar]
- Vudattu NK, Holler E, Ewing P, et al. Reverse signaling of membrane-integrated tumour necrosis factor differentially regulates alloresponses of CD4+ and CD8+ T cells against human microvascular endothelial cells. Immunology 2005; 115 : 536–43. [Google Scholar]
- Van den Brande JM, Braat H, van den Brink GR, et al. Infliximab but not etanercept induces apoptosis in lamina propria T-lymphocytes from patients with Crohn’s disease. Gastroenterology 2003; 124 : 1774–85. [Google Scholar]
- Scallon BJ, Moore MA, Trinh H, et al. Chimeric anti-TNF-alpha monoclonal antibody cA2 binds recombinant transmembrane TNF-alpha and activates immune effector functions. Cytokine 1995; 7 : 251–9. [Google Scholar]
- Ten Hove T, van den Blink B, Pronk I, et al. Dichotomal role of inhibition of p38 MAPK with SB 203580 in experimental colitis. Gut 2002; 50 : 507–12. [Google Scholar]
- Lügering A, Schmidt M, Lügering N, et al. Infliximab induces apoptosis in monocytes from patients with chronic active Crohn’s disease by using a caspase-dependent pathway. Gastroenterology 2001; 121 : 1145–57. [Google Scholar]
- Catrina AI, Trollmo C, Klint E, et al. Evidence that anti-tumor necrosis factor therapy with both etanercept and infliximab induces apoptosis in macrophages, but not lymphocytes, in rheumatoid arthritis joints: extended report. Arthritis Rheum 2005; 52 : 61–72. [Google Scholar]
- Fossati G, Nesbitt AM. In vitro complement-dependent cytotoxicity and antibody-dependent cellular cytotoxicity by the anti-TNF agents adalimumab, etanercept, infliximab and certolizumab pegol (CDO870). Am J Gastroenterol 2005; 100 : S299. [Google Scholar]
- Malaviya R, Sun Y, Tan JK, et al. Etanercept induces apoptosis of dermal dendritic cells in psoriatic plaques of responding patients. J Am Acad Dermatol 2006; 55 : 590–7. [Google Scholar]
- Varfolomeev EE, Ashkenazi A. Tumor necrosis factor: an apoptosis JuNKie ? Cell 2004; 116 : 491–7. [Google Scholar]
- Chaudhary R, Butler M, Playford RJ, Gosh S. Anti-TNF antibody induced stimulated T lymphocyte apoptosis depends on the concentration of the antibody and etanercept induces apoptosis at rates equivalent to infliximab and adalimumab at 10 micrograms per ml concentration. Gastroenterology 2007; 130 : A696. [Google Scholar]
- Anolik JH, Owen T, Barnard J, Sanz I. Anti-tumor necrosis factor therapy in rheumatoid arthritis alters B lymphocyte dynamics. Arthritis Rheum 2005; 52 : S677. [Google Scholar]
- Han S, Zhang X, Marinova E, et al. Blockade of lymphotoxin pathway exacerbates autoimmune arthritis by enhancing the Th1 response. Arthritis Rheum 2005; 52 : 3202–9. [Google Scholar]
- Nestorov I. Clinical pharmacokinetics of TNF antagonists: how do they differ ? Semin Arthritis Rheum 2005; 34 : 12–8. [Google Scholar]
- Zhou H. Clinical pharmacokinetics of etanercept: a fully humanized soluble recombinant tumor necrosis factor receptor fusion protein. J Clin Pharmacol 2005; 45 : 490–7. [Google Scholar]
- Lobo ED, Hansen RJ, Balthasar JP. Antibody pharmacokinetics and pharmacodynamics. J Pharm Sci 2004; 93 : 2645–68. [Google Scholar]
- Kohno T, Louie JS, Stevens SR. Differences in Fc receptor and C1q binding in tumor necrosis factor (TNF) antagonists may contribute to differences in mechanisms of action. J Invest Dermatol 2005; 124 : A111. [Google Scholar]
- Saliu OY, Sofer C, Stein DS, et al. Tumor-necrosis-factor blockers: differential effects on mycobacterial immunity. J Infect Dis 2006; 194 : 486–92. [Google Scholar]
- Kim EY, Priatel JJ, Teh SJ, Teh HS. TNF receptor type 2 (p75) functions as a costimulator for antigen-driven T cell responses in vivo. J Immunol 2006; 176 : 1026–35. [Google Scholar]
- Ehrenstein MR, Evans JG, Singh A, et al. Compromised function of regulatory T cells in rheumatoid arthritis and reversal by anti-TNFalpha therapy. J Exp Med 2004; 200 : 277–85. [Google Scholar]
- Valencia X, Stephens G, Goldbach-Mansky R, et al. TNF downmodulates the function of human CD4+CD25hi T-regulatory cells. Blood 2006; 108 : 253–61. [Google Scholar]
- Hamdi H, Mariette X, Godot V, et al. Inhibition of anti-tuberculosis T-lymphocyte function with tumour necrosis factor antagonists. Arthritis Res Ther 2006; 8 : R114. [Google Scholar]
Liste des tableaux
Caractéristiques des différents anti-TNF. PEG : polyéthylène glycol. TNF sol : tumor necrosis factor soluble ; TNF mb : tumor necrosis factor membranaire ; CDC : complement-dependent cytotoxicity ; ADCC : antibody-dependent cellular cytotoxicity ; LT : lymphotoxine.
Liste des figures
|
Figure 1. Schéma des cinq molécules anti-TNF utilisées en pathologie humaine. PEG : polyéthylène glycol ; CDR : complémentation determining region. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.