")
")
| Issue |
Med Sci (Paris)
Volume 25, Number 11, Novembre 2009
|
|
|---|---|---|
| Page(s) | 931 - 938 | |
| Section | M/S revues | |
| DOI | https://doi.org/10.1051/medsci/20092511931 | |
| Published online | 15 novembre 2009 | |
Thérapies cellulaires pro-angiogéniques dans le traitement des pathologies ischémiques
Proangiogenic cell-based therapy for treatment of ischemic diseases
Paris-Cardiovascular, Research Center-Inserm U970, Hôpital européen Georges Pompidou, 56, rue Leblanc, 75015 Paris, France
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Résumé
L’administration de cellules souches adultes est une option thérapeutique d’intérêt pour les patients présentant une artériopathie des membres inférieurs ou un infarctus du myocarde. L’efficacité de l’injection de différents types de cellules souches/progénitrices adultes autologues dérivées de la moelle osseuse, du sang périphérique ou encore de tissus a été évaluée dans des modèles expérimentaux. Ces cellules, notamment celles d’origine médullaire ou adipocytaire, font l’objet d’essais cliniques. Cette revue synthétise les principaux mécanismes moléculaires et cellulaires sous-tendant l’action des cellules souches/progénitrices adultes sur le compartiment vasculaire et propose quelques approches pour améliorer leur efficacité thérapeutique chez les patients ischémiques.
Abstract
The application of endothelial progenitor cells (EPC) cell-based therapy for regenerative medicine constitutes a promising therapeutic avenue for the treatment of cardiovascular diseases. Based on experimental studies demonstrating that bone marrow-, blood- or tissue-derived stem/progenitor cells improve the functional recovery after ischemia, clinical trials were initiated to address this new therapeutic concept. Although autolougous cell therapy was shown to improve perfusion and function of ischemic tissues, a number of issues remain to be adressed. The nature of the mobilizing, migratory and homing signals, and the mechanisms of action need to be identified and further defined. In addition, strategies to enhance homing, survival and therapeutic potential of EPC need to be developped to improve therapeutic effect and counteract EPC dysfunction in aged patients with cardiovascular risk factors. The present review article will discuss the mechanisms of action of different types of adult stem cells and several approaches to improve their therapeutic efficiency.
© 2009 médecine/sciences - Inserm / SRMS
Les pathologies ischémiques restent une cause importante de morbidité et de mortalité dans les pays industrialisés malgré le développement de nouvelles modalités thérapeutiques comme les approches pharmacologiques ou chirurgicales. Ainsi, l’artériopathie des membres inférieurs est toujours associée à une morbidité élevée et à une baisse de la qualité de vie alors que l’ischémie myocardique, à l’origine d’une insuffisance cardiaque post-infarctus, est source d’une mortalité élevée. À partir des données expérimentales démontrant l’efficacité de l’injection de cellules souches ou progénitrices sur la fonction cardiaque après un infarctus ou sur la perfusion tissulaire dans des modèles d’ischémie périphérique, des études cliniques ont été menées, dès 2001, pour traiter des patients présentant une ischémie cardiaque ou périphérique avec des cellules circulantes, dérivées de la moelle osseuse ou d’autres tissus.
Les différents types de cellules souches adultes
Cellules souches embryonnaires et adultes
D’un point de vue sémantique, une cellule souche est une cellule indifférenciée, capable de s’autorenouveler et de se différencier en des types cellulaires variés. Il existe plusieurs types de cellules souches :
des cellules souches multipotentes capables de donner naissance à plusieurs types cellulaires : par exemple les cellules souches hématopoïétiques de la moelle osseuse qui sont à l’origine des cellules sanguines ;
des cellules souches pluripotentes, issues d’un embryon de 5 à 7 jours, capables de donner naissance à plus de 200 types cellulaires représentatifs de tous les tissus de l’organisme ;
des cellules souches totipotentes embryonnaires capables de donner naissance à tous les types de cellules de l’organisme et les seules capables d’assurer le développement complet d’un individu.
Pour la médecine régénérative, deux principaux types de cellules souches sont à considérer : les cellules souches embryonnaires et les cellules souches adultes. Il existe également des cellules souches fœtales dans le sang de cordon ombilical. Enfin, il convient d’ajouter une quatrième catégorie puisqu’a été récemment révélée la possibilité d’obtenir, à partir notamment de fibroblastes cutanés, des cellules souches phénotypiquement semblables aux cellules souches embryonnaires : les cellules souches pluripotentes induites (iPS, induced pluripotent stem cells) [1]. Alors que les cellules souches embryonnaires ou les cellules souches pluripotentes induites possèdent une réelle capacité à se différencier en plusieurs types de cellules matures appartenant à des organes de natures distinctes, la plasticité cellulaire de la plupart des cellules souches adultes est moindre et celles-ci expriment probablement un phénotype déjà plus différencié. Enfin, il est très probable que ces cellules souches adultes constituent un assemblage hétérogène de différents types cellulaires : non seulement immatures mais également déjà engagés dans un processus de différenciation. Leur utilisation dans le processus de régénération tissulaire apparaît donc plus limitée. Néanmoins, nous focaliserons cette revue sur les différents types de cellules souches adultes et sur leur efficacité thérapeutique dans l’ischémie tissulaire ou dans le traitement de l’infarctus du myocarde.
Cellules souches adultes utilisées en thérapie cellulaire
Les cellules souches adultes se répartissent en au moins trois groupes distincts : les cellules souches dérivées de la moelle osseuse, le pool circulant de cellules souches ou progénitrices qui dérivent, au moins en partie, de la moelle osseuse, et les cellules souches résidentes tissulaires.
Les cellules souches dérivées de la moelle osseuse sont les mieux caractérisées et ont été utilisées dans la plupart des essais cliniques. La moelle osseuse contient un assortiment complexe de cellules souches/progénitrices parmi lesquelles les cellules souches hématopoïétiques, les cellules souches mésenchymateuses, une population de cellules définie par leur exclusion du colorant Hoechst et appelée side population1, les cellules stromales et les cellules progénitrices multipotentes adultes2. Plusieurs études ont étayé le potentiel thérapeutique de l’administration de chacun de ces types cellulaires ; certaines ont aussi montré l’intérêt de l’administration des cellules mononucléées totales de la moelle osseuse dans la survie du tissu ischémique [2].
Progéniteurs endothéliaux circulants
Les cellules progénitrices circulantes représentent la seconde population de cellules progénitrices dont le potentiel thérapeutique a été analysé. Asahara et Isner ont été les premiers à identifier des cellules progénitrices endothéliales (CPE) définies par leur capacité à se différencier en cellules endothéliales, à former des nouveaux capillaires et à initier la revascularisation post-ischémique. Leur capacité de prolifération élevée les distingue des cellules endothéliales matures détachées de la paroi vasculaire [3]. Ces CPE sont caractérisées classiquement par l’expression d’au moins deux marqueurs de cellules souches/progéniteurs hématopoïétiques (CD133 et CD34) et d’un marqueur de cellules endothéliales : le récepteur du VEGF (vascular endothelial growth factor) de type 2 (ou KDR ou flk-1). La sous-population de CPE CD133+VEGFR2+ mais dépourvue de l’antigène CD34, représenterait des précurseurs en amont des CPE CD34+CD133+ et exprimerait un potentiel de régénération vasculaire marqué [4]. Cependant, l’identification des CPE reste un problème majeur. Il a ainsi été suggéré que la sous-population CD34+CD45-, et non celle qui coexprime aussi CD133 et VEGFR2 (CD34+CD133+VEGFR2+), pourrait acquérir un phénotype endothélial [5]. De plus, les cellules myéloides CD14+ peuvent aussi acquérir des caractéristiques de cellules CPE [6, 7]. Enfin, les cultures de cellules mononucléées de sang périphérique révèlent deux types de CPE : les CPE précoces et les CPE tardives. Les CPE précoces ont des caractéristiques de cellules inflammatoires et induisent la formation des tubes vasculaires indirectement, par une voie paracrine. Les CPE tardives expriment les marqueurs caractéristiques des cellules endothéliales, comme le récepteur du VEGF de type 2 et la VE-Cadhérine, et se distinguent par leur réel potentiel de différenciation en cellules endothéliales capables de s’incorporer dans les capillaires néoformés. Il est donc probable que ce terme de CPE désigne des cellules circulantes hétérogènes provenant de multiples précurseurs et correspondant à des stades de maturation différents de cellules endothéliales. L’absence de marqueurs définissant spécifiquement la population de CPE productrice de cellules endothéliales fonctionnelles est préjudiciable à une dissection plus précise de ces populations. Il est également plausible que différents types de cellules souches aient un potentiel pro-angiogénique qu’elles expriment via des mécanismes et voies de différenciation divers. Ces deux mécanismes pro-angiogéniques semblent néanmoins acquis : d’une part des cellules progénitrices stimulent la néovascularisation par leur effet paracrine et d’autre part, des CPE « réelles » assurent la formation de néovaisseaux par leur réelle capacité à se différencier en cellules endothéliales.
Autres populations de progéniteurs tissulaires
Les autres populations de cellules souches/progéniteurs ayant un potentiel thérapeutique dans un contexte ischémique dérivent essentiellement des cellules stromales du tissu adipeux (CPE et cellules souches mésenchymateuses) ou des cellules souches du tissu cardiaque. Le tissu adipeux est une source de différentes sous-populations de cellules souches/progénitrices utiles à la régénération cardiaque et à l’augmentation de la néovascularisation. Des cellules souches mésenchymateuses et des CPE ont été isolées après digestion enzymatique du tissu adipeux et leurs effets bénéfiques ont été démontrés dans des modèles expérimentaux [8]. Plus récemment, plusieurs laboratoires ont isolé des cellules progénitrices dans le tissu cardiaque chez des rongeurs : il s’agit notamment de cellules exprimant c-kit, Sca-1, Islet-1 et ou se projetant dans la side population après marquage au Hoechst 33342. Ces cellules souches sont multipotentes in vitro et se différencient in vivo en cardiomyocytes et cellules vasculaires coronaires [9]. Même si ces travaux ont été bien évidemment rapidement remis en question [10], la présence de cellules souches résidant dans le tissu cardiaque représente une avancée majeure dans le domaine de la cardiologie, d’un point de vue biologique mais également sur le plan clinique. Elle permettrait notamment de comprendre l’observation d’un nombre restreint de cardiomyocytes en mitose et la présence de cellules progénitrices vasculaires dans le tissu cardiaque, dont l’origine, médullaire ou endogène cardiaque, est très débattue. Enfin, ces cellules souches cardiaques pourraient servir de base à des thérapies cellulaires développées pour le traitement des pathologies cardiaques [11].
Mécanisme d’action des cellules souches adultes
En réponse à une ischémie tissulaire, les CPE d’origines médullaire, tissulaire ou périphérique migrent vers le tissu ischémique (Figure 1). Le recrutement des CPE exprimant CXCR4 est stimulé par l’hypoxie et le facteur de transcription HIF-1α qui active l’expression du ligand de CXCR4, la chimiokine SDF-1/CCL12 [12]. Les cellules mononucléées ou les CPE provenant d’animaux Cxcr4+/− n’ayant qu’un allèle codant pour CXCR4 expriment un nombre diminué de récepteurs CXCR4 et leur capacité à stimuler la reperfusion tissulaire chez des souris immunodéficientes s’en trouve affaiblie [13]. L’hypoxie locale libère également des facteurs solubles chimio-attractants pour les CPE circulantes. Le VEGF est un candidat potentiel dont le taux augmente en conditions hypoxiques et qui induit SDF-1 dans les structures vasculaires et périvasculaires [14]. Les chimiokines et les plaquettes permettent aussi la capture des CPE au niveau des zones de lésions vasculaires [15, 16]. Une fois qu’elles ont été recrutées, les CPE se fixent sur l’endothélium activé par l’ischémie, grâce à l’action des molécules d’adhésion présentes sur la surface vasculaire comme l’intégrine β2 et la sélectine de type L [17–19]. Une fois adhérentes à l’endothélium activé par l’ischémie, les CPE exercent leurs effets bénéfiques sur la néogenèse vasculaire par des mécanismes variés (Figure 2).
 |
Figure 1. Mécanimes simplifiés d’action des cellules souches de la moelle osseuse dans la réparation vasculaire. Classiquement, il est admis que des signaux provenant du tissu ischémique entraînent la mobilisation de cellules souches de la moelle osseuse vers le sang où elles se différencient en cellules progénitrices endothéliales (CPE). Celles-ci migrent vers la zone lésée et s’arrêtent au niveau de l’endothélium activé par l’ischémie où elles vont exercer leur potentiel pro-angiogénique soit par une action paracrine : libération de monoxyde d’azote (NO), de facteurs de croissance (FC) ou de molécules anti-inflammatoires (comme l’interleukine-10), soit par leur aptitude à se différencier en cellules endothéliales et à s’incorporer dans les néocapillaires. PSGL1 : P selectin glycoprotein ligand 1. |
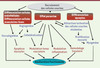 |
Figure 2. Mécanismes à l’origine de l’efficacité thérapeutique des cellules souches. Ils reposent sur des mécanismes moléculaires et cellulaires complexes : potentiel de différenciation soit en cellules vasculaires ou myocytaires et activité paracrine. Il en résulte une modulation quantitative (vasculogenèse et angiogenèse) et qualitative (collatéralisation et inflexion du tonus vasculaire) du compartiment vasculaire mais également myocytaire. |
Selon une première hypothèse, les progéniteurs s’incorporaient dans le site de néovascularisation où ils activent leur différenciation en cellules endothéliales matures [20]. Cependant, la contribution des CPE à la néogenèse vasculaire est très variable selon le modèle expérimental, le type et le sous-type de progéniteurs ou encore la méthode d’isolement des CPE. Certains travaux expérimentaux suggèrent même que ces progéniteurs ne s’incorporent pas dans les néocapillaires mais interviennent surtout par leur rôle de cellules de soutien : les progéniteurs d’origine médullaire s’accumulent autour des vaisseaux collatéraux et se différencient en fibroblastes, leucocytes et péricytes source de facteurs de croissance et de chimiokines [21]. La libération de SDF-1 par les plaquettes favorise la mobilisation et le recrutement des progéniteurs CXCR4+VEGFR1+ autour des structures vasculaires qui, à leur tour, promeuvent le développement de vaisseaux collatéraux [22]. Ainsi, le rôle premier des progéniteurs pourrait être de libérer des facteurs angiogéniques et artériogéniques contribuant ainsi à la régénération vasculaire par des effets paracrines. Par leur capacité à réguler le tonus vasculaire et à induire une vasodilatation via un mécanisme dépendant de la voie eNOS (endothelial nitric oxide synthase), les cellules progénitrices sont également susceptibles de participer à l’amélioration de la perfusion tissulaire dans le territoire ischémié [23]. Enfin, les cellules progénitrices, par leur sécrétion de cytokines anti-inflammatoires comme l’interleukine-10, peuvent améliorer la fonction cardiaque après un infarctus du myocarde [24].
Cependant, les effets paracrines et le potentiel de différenciation en cellules vasculaires ou cardiaques des cellules souches/progéniteurs adultes ne doivent pas être opposés, mais plutôt considérés comme complémentaires. Ainsi, dans un modèle d’ischémie des membres inférieurs et du myocarde chez la souris nude greffée avec des cellules humaines pro-angiogéniques transduites avec un gène « suicide » (séquence du gène TK, thymidine kinase, du virus herpes simplex de type 2), l’induction de la mort des cellules progénitrices par le ganciclovir 14 jours après leur injection annule le bénéfice thérapeutique sur la fonction cardiaque. Cela révèle ainsi que l’incorporation physique et la persistance des cellules contribuent à l’effet de ces dernières sur la vascularisation et la fonction cardiaque [25] et conforte l’hypothèse d’une complémentarité des mécanismes paracrines et intrinsèques.
Applications cliniques
Ces thérapies cellulaires sont actuellement testées dans des essais cliniques incluant de petites cohortes de patients. La moelle osseuse est, pour l’instant, la principale source de cellules souches adultes à la base des thérapies cellulaires pro-angiogéniques en situation autologue. La transition rapide de la démarche expérimentale à l’application clinique a été facilitée par plus de trente ans d’expérience clinique dans le domaine des transplantations de moelle osseuse et par l’absence d’effet délétère de ces transplantations pour la reconstitution du système hématopoïétique. Certes ces cellules médullaires sont un mélange de cellules matures et immatures avec les avantages et inconvénients propres à chacune de ces catégories, mais leur facilité d’obtention permet d’envisager leur utilisation en situation aiguë. Les autres types de cellules souches adultes requièrent une étape d’amplification ex vivo, ce qui impose un délai entre le moment du prélèvement et l’administration des cellules. Une alternative serait de disposer d’une banque de cellules autologues, une autre de greffer des cellules allogéniques.
L’implantation autologue de cellules mononucléées médullaires dans le membre ischémié de patients souffrant d’une maladie artérielle périphérique ou de thromboangiopathie oblitérante améliore la vascularisation et le processus cicatriciel [26, 27]. Cependant, un certain nombre de ces patients présentent des effets secondaires, et l’amélioration n’est souvent que transitoire [27]. De même, la transplantation par voie intracoronaire de cellules mononucléées médullaires dans le myocarde de patients ayant une ischémie aiguë augmente la perfusion et la fonction du tissu ischémié [28–30]. Dans cette indication également, certaines études montrent le caractère transitoire de l’amélioration, d’autres une absence d’effet, soulignant ainsi la nécessité de poursuivre l’analyse à la fois expérimentale et clinique de l’efficacité thérapeutique de ces stratégies [31, 32].
Quelles stratégies pour optimiser les thérapies cellulaires pro-angiogéniques ?
Des études récentes ont démontré que la plupart des facteurs de risque cardiovasculaire comme le diabète, l’hypertension ou l’âge affectent la fonction des cellules souches adultes [33, 34]. Cette altération des cellules souches du patient susceptibles d’être utilisées pour une transplantation autologue incite à réfléchir à des stratégies visant à restaurer leur activité afin de ne pas compromettre leur efficacité thérapeutique (Figure 3). Elles pourraient être de plusieurs types.
 |
Figure 3. Altération fonctionnelle des cellules souches chez les patients à risque cardiovasculaire et stratégies d’amélioration. La plupart des facteurs de risque cardiovasculaire inhibent la fonction des cellules souches/progénitrices. Dans le cas de la transplantation autologue de cellules souches adultes pour le traitement des patients ischémiques, un certain nombre de stratégies peuvent être envisagées pour restaurer la fonction de ces cellules : transfert de gènes stimulant la prolifération, prétraitement par des molécules pharmacologiques ou encore co-traitements utilisant l’Ephrine B2 ou différents types de cellules progénitrices : dnGSK3β, dominant négatif de la protéine kinase GSK3β ; TERT, télomérase transcriptase inverse ; IEC, inhibiteur de l’enzyme de conversion, SDF-1, facteur dérivé du stroma (ligand de CXCR4), CPML, cellules progénitrices musculaires lisses. |
Modifications génétiques
Ainsi, l’inhibition de la GSK3β (glycogen synthase kinase 3) augmente la sécrétion de VEGF et d’interleukine-8, la survie et la prolifération des CPE précoces et la survie et la différenciation des CPE tardives [35]. Le transfert de gènes codant pour le VEGFA164 ou pour la sous-unité catalytique de la télomérase Tert (telomerase reverse transcriptase) augmente également les activités mitogénique et migratoire et le potentiel thérapeutique des CPE dans un modèle d’ischémie du membre inférieur chez la souris immunodéficiente [36, 37]. Enfin, le transfert du gène codant pour HIF-1α est accompagné d’une augmentation d’expression du VEGF et de son récepteur VEGFR2. Il en résulte in vivo une plus grande efficacité des CPE transfectées sur le processus de revascularisation post-ischémique [38].
Associer thérapie cellulaire et stratégies pro-angiogéniques
Des thérapies combinant la transplantation de CPE avec des stratégies pro-angiogéniques pourraient corriger le déficit fonctionnel des CPE chez les patients ischémiques. L’utilisation de molécules pharmacologiques - les statines par exemple - inhibe la sénescence des CPE et induit leur prolifération [39]. Les inhibiteurs de l’enzyme de conversion (IEC) activent la néogenèse vasculaire en stimulant la voie dépendante de la bradykinine [40]. Le traitement de patients coronariens par l’IEC Ramipril augmente le nombre de CPE circulantes et leur activité biologique [41]. Un effet additif a même été observé entre l’administration intraveineuse de CPE et celle d’un IEC sur la fonction ventriculaire de rats immunodéficients chez lesquels un infarctus du myocarde avait été induit [42]. Enfin, l’utilisation d’inhibiteurs du stress oxydant restaure la capacité de cellules mononucléées médullaires de souris diabétiques ou de rats hypertendus à se différencier en cellules endothéliales et à démarrer un processus de néogenèse vasculaire dans la zone ischémiée [43, 44].
Associer thérapies génique et cellulaire
Cette combinaison présente un intérêt. Ainsi, l’injection locale de SDF-1 dans la patte ischémique de souris immunodéficientes associée à l’administration intraveineuse de CPE humaines augmente l’accumulation tissulaire de CPE dans la zone lésée et leur efficacité thérapeutique [45]. Un prétraitement par l’éphrine B2 améliore également la capacité de migration des CPE vers la zone ischémique et ainsi leur potentiel thérapeutique dans un modèle d’ischémie du membre inférieur [46]. De même, l’administration simultanée de CPE et de progéniteurs de cellules musculaires lisses est bénéfique car la libération de l’angiopoïétine-2 par ces derniers active la survie des CPE dans le territoire ischémique et ainsi facilite la néovascularisation [47].
Conclusion
Il apparaît donc clairement que l’administration de cellules souches ou progénitrices adultes pour la régénération tissulaire est encore loin d’être une stratégie de routine en pratique clinique. Une attention toute particulière doit être apportée aux types de cellules souches utilisées, à leur origine, leur pureté, leur capacité de différenciation et à leur potentiel thérapeutique, en particulier si elles sont utilisées pour une transplantation autologue chez des patients ischémiques déjà porteurs de facteurs de risque cardiovasculaire importants. L’avenir de la thérapie cellulaire pro-angiogénique réside peut-être dans la transplantation allogénique de cellules souches provenant de sang de cordon ombilical. Mais se pose le problème de la compatibilité immunologique et du développement des banques de sang de cordon, notamment en France. Enfin, même si l’utilisation de cellules souches embryonnaires pourrait, en partie, résoudre les difficultés fonctionnelles inhérentes aux cellules souches adultes, leur potentiel thérapeutique doit être évalué au regard des risques inhérents à leur utilisation dans les tissus pathologiques chez les patients ischémiques [48].
Cette appellation vient de l’analyse au cytomètre de flux de cellules de moelle osseuse ayant été incubées avec le colorant Hoechst 33342 : lorsque l’émission de fluorescence est analysée simultanément à deux longueurs d’onde après excitation du Hoechst dans l’UV, on distingue une population minoritaire se distribuant comme une « queue » expulsant le colorant et donc émettant une faible fluoresence, à la différence de la majorité des cellules. Elle a été nommée side population ou SP.
On désigne sous ce terme une population de cellules très rares, dont l’existence est encore controversée, et qui, in vitro, expriment un potentiel de différenciation très large, y compris dans des lignées non mésodermiques.
Références
- Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006; 126 : 663–76. [Google Scholar]
- Dimmeler S, Burchfield J, Zeiher AM. Cell-based therapy of myocardial infarction. Arterioscler Thromb Vasc Biol 2008; 28 : 208–16. [Google Scholar]
- Rafii S, Lyden D. Therapeutic stem and progenitor cell transplantation for organ vascularization and regeneration. Nat Med 2003; 9 : 702–12. [Google Scholar]
- Friedrich EB, Walenta K, Scharlau J, et al. CD34-/CD133+/VEGFR-2+ endothelial progenitor cell subpopulation with potent vasoregenerative capacities. Circ Res 2006; 98 : e20–25. [Google Scholar]
- Case J, Mead LE, Bessler WK, et al. Human CD34+AC133+VEGFR-2+ cells are not endothelial progenitor cells but distinct, primitive hematopoietic progenitors. Exp Hematol 2007; 35 : 1109–18. [Google Scholar]
- Urbich C, Heeschen C, Aicher A, et al. Relevance of monocytic features for neovascularization capacity of circulating endothelial progenitor cells. Circulation 2003; 108 : 2511–6. [Google Scholar]
- Rehman J, Li J, Orschell CM, et al. Peripheral blood endothelial progenitor cells are derived from monocyte/macrophages and secrete angiogenic growth factors. Circulation 2003; 107 : 1164–9. [Google Scholar]
- Planat-Benard V, Silvestre JS, Cousin B, et al. Plasticity of human adipose lineage cells toward endothelial cells: physiological and therapeutic perspectives. Circulation 2004; 109 : 656–63. [Google Scholar]
- Beltrami AP, Barlucchi L, Torella D, et al. Adult cardiac stem cells are multipotent and support myocardial regeneration. Cell 2003; 114 : 763–76. [Google Scholar]
- Laflamme MA, Murry CE. Regenerating the heart. Nat Biotechnol 2005; 23 : 845–56. [Google Scholar]
- Dawn B, Stein AB, Urbanek K, et al. Cardiac stem cells delivered intravascularly traverse the vessel barrier, regenerate infarcted myocardium, and improve cardiac function. Proc Natl Acad Sci USA 2005; 102 : 3766–71. [Google Scholar]
- Ceradini DJ, Gurtner GC. Homing to hypoxia: HIF-1 as a mediator of progenitor cell recruitment to injured tissue. Trends Cardiovasc Med 2005; 15 : 57–63. [Google Scholar]
- Walter DH, Haendeler J, Reinhold J, et al. Impaired CXCR4 signaling contributes to the reduced neovascularization capacity of endothelial progenitor cells from patients with coronary artery disease. Circ Res 2005; 97 : 1142–51. [Google Scholar]
- Grunewald M, Avraham I, Dor Y, et al. VEGF-induced adult neovascularization: recruitment, retention, and role of accessory cells. Cell 2006; 124 : 175–89. [Google Scholar]
- Waeckel L, Mallat Z, Potteaux S, et al. Impairment in postischemic neovascularization in mice lacking the CXC chemokine receptor 3. Circ Res 2005; 96 : 576–82. [Google Scholar]
- Langer H, May AE, Daub K, et al. Adherent platelets recruit and induce differentiation of murine embryonic endothelial progenitor cells to mature endothelial cells in vitro. Circ Res 2006; 98 : e2–10. [Google Scholar]
- Chavakis E, Aicher A, Heeschen C, et al. Role of beta2-integrins for homing and neovascularization capacity of endothelial progenitor cells. J Exp Med 2005; 201 : 63–72. [Google Scholar]
- Biancone L, Cantaluppi V, Duo D, et al. Role of L-selectin in the vascular homing of peripheral blood-derived endothelial progenitor cells. J Immunol 2004; 173 : 5268–74. [Google Scholar]
- Yoon CH, Hur J, Oh IY, et al. Intercellular adhesion molecule-1 is upregulated in ischemic muscle, which mediates trafficking of endothelial progenitor cells. Arterioscler Thromb Vasc Biol 2006; 26 : 1066–72. [Google Scholar]
- Takahashi T, Kalka C, Masuda H, et al. Ischemia- and cytokine-induced mobilization of bone marrow-derived endothelial progenitor cells for neovascularization. Nat Med 1999; 5 : 434–8. [Google Scholar]
- Ziegelhoeffer T, Fernandez B, Kostin S, et al. Bone marrow-derived cells do not incorporate into the adult growing vasculature. Circ Res 2004; 94 : 230–8. [Google Scholar]
- Jin DK, Shido K, Kopp HG, et al. Cytokine-mediated deployment of SDF-1 induces revascularization through recruitment of CXCR4+ hemangiocytes. Nat Med 2006; 12 : 557–67. [Google Scholar]
- You D, Waeckel L, Ebrahimian TG, et al. Increase in vascular permeability and vasodilation are critical for proangiogenic effects of stem cell therapy. Circulation 2006; 114 : 328–38. [Google Scholar]
- Burchfield JS, Iwasaki M, Koyanagi M, et al. Interleukin-10 from transplanted bone marrow mononuclear cells contributes to cardiac protection after myocardial infarction. Circ Res 2008; 103 : 203–11. [Google Scholar]
- Ziebart T, Yoon CH, Trepels T, et al. Sustained persistence of transplanted proangiogenic cells contributes to neovascularization and cardiac function after ischemia. Circ Res 2008; 103 : 1327–34. [Google Scholar]
- Tateishi-Yuyama E, Matsubara H, Murohara T, et al. Therapeutic angiogenesis for patients with limb ischaemia by autologous transplantation of bone-marrow cells: a pilot study and a randomised controlled trial. Lancet 2002; 360 : 427–35. [Google Scholar]
- Miyamoto K, Nishigami K, Nagaya N, et al. Unblinded pilot study of autologous transplantation of bone marrow mononuclear cells in patients with thromboangiitis obliterans. Circulation 2006; 114 : 2679–84. [Google Scholar]
- Schachinger V, Assmus B, Britten MB, et al. Transplantation of progenitor cells and regeneration enhancement in acute myocardial infarction: final one-year results of the TOPCARE-AMI Trial. J Am Coll Cardiol 2004; 44 : 1690–9. [Google Scholar]
- Assmus B, Honold J, Schachinger V, et al. Transcoronary transplantation of progenitor cells after myocardial infarction. N Engl J Med 2006; 355 : 1222–32. [Google Scholar]
- Schachinger V, Erbs S, Elsasser A, et al. Intracoronary bone marrow-derived progenitor cells in acute myocardial infarction. N Engl J Med 2006; 355 : 1210–21. [Google Scholar]
- Lunde K, Solheim S, Aakhus S, et al. Intracoronary injection of mononuclear bone marrow cells in acute myocardial infarction. N Engl J Med 2006; 355 : 1199–209. [Google Scholar]
- Meyer GP, Wollert KC, Lotz J, et al. Intracoronary bone marrow cell transfer after myocardial infarction: eighteen months’ follow-up data from the randomized, controlled BOOST (Bone marrow transfer to enhance ST-elevation infarct regeneration) trial. Circulation 2006; 113 : 1287–94. [Google Scholar]
- You D, Cochain C, Loinard C, et al. Hypertension impairs postnatal vasculogenesis: role of antihypertensive agents. Hypertension 2008; 51 : 1537–44. [Google Scholar]
- Tamarat R, Silvestre JS, Le Ricousse-Roussanne S, et al. Impairment in ischemia-induced neovascularization in diabetes: bone marrow mononuclear cell dysfunction and therapeutic potential of placenta growth factor treatment. Am J Pathol 2004; 164 : 457–66. [Google Scholar]
- Choi JH, Hur J, Yoon CH, et al. Augmentation of therapeutic angiogenesis using genetically modified human endothelial progenitor cells with altered glycogen synthase kinase-3beta activity. J Biol Chem 2004; 279 : 49430–8. [Google Scholar]
- Iwaguro H, Yamaguchi J, Kalka C, et al. Endothelial progenitor cell vascular endothelial growth factor gene transfer for vascular regeneration. Circulation 2002; 105 : 732–8. [Google Scholar]
- Murasawa S, Llevadot J, Silver M, et al. Constitutive human telomerase reverse transcriptase expression enhances regenerative properties of endothelial progenitor cells. Circulation 2002; 106 : 1133–9. [Google Scholar]
- Jiang M, Wang B, Wang C, et al. Angiogenesis by transplantation of HIF-1 alpha modified EPCs into ischemic limbs. J Cell Biochem 2008; 103 : 321–34. [Google Scholar]
- Assmus B, Urbich C, Aicher A, et al. HMG-CoA reductase inhibitors reduce senescence and increase proliferation of endothelial progenitor cells via regulation of cell cycle regulatory genes. Circ Res 2003; 92 : 1049–55. [Google Scholar]
- Silvestre JS, Bergaya S, Tamarat R, et al. Proangiogenic effect of angiotensin-converting enzyme inhibition is mediated by the bradykinin B(2) receptor pathway. Circ Res 2001; 89 : 678–83. [Google Scholar]
- Min TQ, Zhu CJ, Xiang WX, et al. Improvement in endothelial progenitor cells from peripheral blood by ramipril therapy in patients with stable coronary artery disease. Cardiovasc Drugs Ther 2004; 18 : 203–9. [Google Scholar]
- Boyle AJ, Schuster M, Witkowski P, et al. Additive effects of endothelial progenitor cells combined with ACE inhibition and beta-blockade on left ventricular function following acute myocardial infarction. J Renin Angiotensin Aldosterone Syst 2005; 6 : 33–7. [Google Scholar]
- Ebrahimian TG, Heymes C, You D, et al. NADPH oxidase-derived overproduction of reactive oxygen species impairs postischemic neovascularization in mice with type 1 diabetes. Am J Pathol 2006; 169 : 719–28. [Google Scholar]
- You D, Cochain C, Loinard C, et al. Combination of the angiotensin converting enzyme inhibitor Perindopril and the diuretic Indapamide activate post-natal vasculogenesis in Spontaneaously Hypertensive Rats. J Pharmacol Exp Ther 2008; 325 : 766–73. [Google Scholar]
- Yamaguchi J, Kusano KF, Masuo O, et al. Stromal cell-derived factor-1 effects on ex vivo expanded endothelial progenitor cell recruitment for ischemic neovascularization. Circulation 2003; 107 : 1322–8. [Google Scholar]
- Foubert P, Silvestre JS, Souttou B, et al. PSGL-1-mediated activation of EphB4 increases the proangiogenic potential of endothelial progenitor cells. J Clin Invest 2007; 117 : 1527–37. [Google Scholar]
- Foubert P, Matrone G, Souttou B, et al. Coadministration of endothelial and smooth muscle progenitor cells enhances the efficiency of proangiogenic cell-based therapy. Circ Res 2008; 103 : 751–60. [Google Scholar]
- Peters BA, Diaz LA, Polyak K, et al. Contribution of bone marrow-derived endothelial cells to human tumor vasculature. Nat Med 2005; 11 : 261–2. [Google Scholar]
Liste des figures
|
Figure 1. Mécanimes simplifiés d’action des cellules souches de la moelle osseuse dans la réparation vasculaire. Classiquement, il est admis que des signaux provenant du tissu ischémique entraînent la mobilisation de cellules souches de la moelle osseuse vers le sang où elles se différencient en cellules progénitrices endothéliales (CPE). Celles-ci migrent vers la zone lésée et s’arrêtent au niveau de l’endothélium activé par l’ischémie où elles vont exercer leur potentiel pro-angiogénique soit par une action paracrine : libération de monoxyde d’azote (NO), de facteurs de croissance (FC) ou de molécules anti-inflammatoires (comme l’interleukine-10), soit par leur aptitude à se différencier en cellules endothéliales et à s’incorporer dans les néocapillaires. PSGL1 : P selectin glycoprotein ligand 1. |
| Dans le texte | |
|
Figure 2. Mécanismes à l’origine de l’efficacité thérapeutique des cellules souches. Ils reposent sur des mécanismes moléculaires et cellulaires complexes : potentiel de différenciation soit en cellules vasculaires ou myocytaires et activité paracrine. Il en résulte une modulation quantitative (vasculogenèse et angiogenèse) et qualitative (collatéralisation et inflexion du tonus vasculaire) du compartiment vasculaire mais également myocytaire. |
| Dans le texte | |
|
Figure 3. Altération fonctionnelle des cellules souches chez les patients à risque cardiovasculaire et stratégies d’amélioration. La plupart des facteurs de risque cardiovasculaire inhibent la fonction des cellules souches/progénitrices. Dans le cas de la transplantation autologue de cellules souches adultes pour le traitement des patients ischémiques, un certain nombre de stratégies peuvent être envisagées pour restaurer la fonction de ces cellules : transfert de gènes stimulant la prolifération, prétraitement par des molécules pharmacologiques ou encore co-traitements utilisant l’Ephrine B2 ou différents types de cellules progénitrices : dnGSK3β, dominant négatif de la protéine kinase GSK3β ; TERT, télomérase transcriptase inverse ; IEC, inhibiteur de l’enzyme de conversion, SDF-1, facteur dérivé du stroma (ligand de CXCR4), CPML, cellules progénitrices musculaires lisses. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.