")
")
| Issue |
Med Sci (Paris)
Volume 25, Number 4, Avril 2009
|
|
|---|---|---|
| Page(s) | 333 - 335 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/2009254333 | |
| Published online | 15 avril 2009 | |
Des cellules souches embryonnaires humaines pour la thérapie cellulaire de la maladie de Huntington
Human embryonic stem cell derived striatal graft for Huntington’s disease cell therapy
Inserm/UEVE UMR-861, I-STEM, AFM, Institut des cellules Souches pour le Traitement et l’Étude des maladies Monogéniques, 5, rue Henri Desbruères, 91030 Évry Cedex, France
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Limitations actuelles d’une thérapie cellulaire de la maladie de Huntington
Comment passer d’une thérapie cellulaire expérimentale qui donne une preuve de concept à une thérapeutique offerte à tous les patients qui pourraient en bénéficier, voilà la question à laquelle nous nous sommes attaqués en identifiant les conditions dans lesquelles des cellules dérivées de lignées de cellules souches embryonnaires (CSE) pourraient être substituées aux précurseurs neuraux prélevés chez des fœtus issus d’IVG (interruption volontaire de grossesse) que l’on greffe aujourd’hui chez des patients atteints de la maladie de Huntington. La maladie de Huntington (MH) est une maladie neurodégénérative d’origine génétique. Elle se caractérise par une atteinte préférentielle des neurones GABAergiques moyens épineux (MSN) du striatum, une structure cérébrale impliquée dans la régulation des fonctions motrices, cognitives et comportementales. La MH se déclare généralement entre 30 et 50 ans et conduit inéluctablement à la mort des patients en 10 à 20 ans. S’il n’existe encore à ce jour aucun traitement curatif de cette pathologie, une stratégie de thérapie cellulaire qui repose sur la transplantation intracérébrale de neuroblastes fœtaux issus du télencéphale ventral (région du cerveau en développement à l’origine du striatum) a permis, depuis une dizaine d’années, d’obtenir des améliorations cliniques notables lors d’essais cliniques impliquant à ce jour plusieurs dizaines de patients [1, 2]. Malgré des résultats encourageants, l’extension de cette thérapie substitutive se heurte à des obstacles aujourd’hui incontournables car inhérents à l’accès même aux cellules chez des donneurs que l’on ne peut atteindre qu’au prix d’une logistique très lourde, rétive à la standardisation et limitante en ce qui concerne les possibilités de contrôle de la qualité des produits. La mise en œuvre à grande échelle de la thérapie cellulaire de la maladie de Huntington nécessite donc l’identification d’une nouvelle source cellulaire permettant de s’affranchir de ces problèmes liés aux tissus fœtaux, tout en procurant des résultats au moins aussi efficaces que ces derniers. Comme pour plusieurs autres indications dans des maladies dégénératives touchant des organes divers, les cellules souches embryonnaires humaines (CSEh) apparaissent ici comme des candidates particulièrement intéressantes du fait de leurs deux propriétés cardinales, l’autorenouvellement - qui assure l’obtention d’une biomasse suffisante, quelle qu’elle soit - et la pluripotence, qui donne accès par différenciation in vitro à n’importe quel phénotype cellulaire [3, 9]. La maîtrise de ces deux propriétés est l’enjeu de toutes les recherches entreprises aujourd’hui pour amener les CSEh aux patients, et notre récente étude [4] démontre que cela est envisageable dans le contexte d’une approche de thérapie substitutive expérimentale de la MH.
Différenciation des CSEh en neurones du striatum in vitro
La première étape en vue de cette utilisation thérapeutique des CSEh est l’élaboration d’un protocole permettant d’orienter la différenciation de ces cellules vers un phénotype pertinent, dans notre cas celui des neurones striataux. Le protocole de différenciation striatale in vitro des CSEh repose sur la réplication d’une succession de trois étapes essentielles de l’ontogenèse : 1- l’induction neurale, 2- l’expansion et la spécification des précurseurs neuraux vers le phénotype striatal, et 3- la différenciation terminale de ces progéniteurs en neurones striataux. La première étape est fondée sur la culture des CSEh avec des cellules stromales de moelle osseuse (MS5) comme cela a déjà été décrit [5]. Cette méthode d’induction neurale permet la production en une vingtaine de jours de cellules neuroépithéliales organisées en structures circulaires caractéristiques, appelées rosettes neurales (Figure 1A). Ces cellules expriment des marqueurs neuraux tels que la protéine associée aux filaments intermédiaires Nestine, ou encore les facteurs de transcription Pax6 et Sox1. Elles sont également caractérisées par l’expression du facteur de transcription BF1, spécifique du télencéphale. La deuxième étape vise à diriger le phénotype de ces cellules neuroépithéliales vers celui de populations cellulaires du télencéphale ventral, ce que nous avons réalisé grâce à deux cytokines, SHH (sonic hedgehog) et DKK1 (dickkopf 1, un inhibiteur de la voie de signalisation Wnt). Au cours de la neurogenèse chez la souris, la voie Shh et la voie Wnt ont des actions antagonistes qui participent respectivement à la ventralisation et à la dorsalisation du télencéphale [6, 7]. L’ajout de Shh et Dkk1 a, de plus, permis la production, à partir de cellules ES de souris, de populations cellulaires enrichies en cellules du télencéphale ventral [8]. Sous l’action combinée des molécules SHH et DKK1, nous avons pu observer une augmentation significative de l’expression des facteurs de transcription spécifiques du télencéphale ventral (GSH2 et DLX2), traduisant l’enrichissement des populations neurales dérivées de CSEh en progéniteurs striataux. La troisième étape, qui aboutit à la différenciation terminale de ces populations cellulaires « enrichies » en progéniteurs striataux, a été obtenue en diminuant la densité cellulaire des culture en présence de BDNF (brain-derived neurotrophic factor), d’AMPc et d’acide valproïque. Cela permet d’engendrer de très nombreux neurones post-mitotiques qui possèdent des caractéristiques phénotypiques de MSN. Après environ 60 jours de culture plus de la moitié des neurones matures (positifs pour protéine MAP2, microtubule-associated protein 2) expriment la protéine DARPP-32 (dopamine- and cAMP-regulated phosphoprotein MR = 32 000), une protéine fortement et spécifiquement exprimée par les MSN (Figure 1B) et plus d’un tiers produisent du GABA (acide gamma-aminobutyrique).
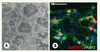 |
Figure 1. Différenciation des cellules souches embryonnaires humaines en neurones striataux. A. Photo en microscopie à contraste de phase d’une colonie de cellules neurales organisées en rosettes après 23 jours de culture sur une monocouche de cellules stromales MS5. B. Après environ 62 jours de différenciation, près de 53% des neurones MAP2+ (en vert) expriment la protéine striatale DARPP32 (en rouge). Barres d’échelle : 100 µm. |
Maturation des populations neuronales dérivées de CSEh in vivo
Nous avons ensuite étudié la survie et la maturation de ces cellules in vivo, après leur transplantation dans le cerveau de rats modèles de la MH. Une première série d’expériences à court terme (4 à 6 semaines) nous a permis de démontrer que la greffe de populations neurales très immatures (différenciées en moins de 21 à 29 jours) a conduit dans de très nombreux cas à la formation de tératomes. À l’inverse, des populations de progéniteurs neuronaux différenciés pendant 45 à 56 jours in vitro procurent les résultats les plus intéressants en termes de survie, de maturation striatale in vivo, et d’absence de formation de tératome. Des expériences à long terme (3 à 5 mois) de xénogreffe de ces progéniteurs ont mis en évidence la présence au sein du greffon de larges zones constituées de neurones humains ayant un phénotype de MSN (DARPP-32+/ HNA+, human nuclear antigen) (Figures 2A et 2B). Ce résultat démontre le fort potentiel de différenciation in vivo des progéniteurs striataux dérivés de CSEh. Cependant, ces expériences ont également révélé un processus de prolifération excessive des cellules implantées qui nécessite l’élaboration de mesures préventives. À partir de deux mois après transplantation, les greffons striataux issus des CSEh occupent in vivo un espace important, très supérieur à celui du striatum, comprimant ainsi les hémisphères cérébraux (Figure 2C). Les analyses histologiques et immunohistochimiques n’ont révélé ni la formation de tératome, ni la présence de CSEh indifférenciées. En revanche, tous les greffons contiennent des régions composées de cellules neurales (Nestine+ et Pax6+) en phase de prolifération (Ki67+) (Figure 2D). Ces régions « prolifératives » et celles contenant des neurones DARPP-32 sont distinctes et n’adoptent pas d’organisation ou de localisation préférentielle. Il ne semble pas que cette activité proliférative qui perdure après transplantation soit liée à des anomalies moléculaires ou fonctionnelles de la descendance des CSEh et les analyses que nous avons conduites suggèrent qu’il s’agit plutôt des conséquences de l’activité proliférative normale des précurseurs neuraux que nous transplantons. En effet, les cellules de la zone germinative de l’éminence ganglionnaire latérale qui donne naissance au striatum fournissent également, dans les mois qui suivent, les interneurones corticaux et des vagues considérables en nombre de cellules gliales, d’astrocytes et d’oligodendrocytes. Il est vraisemblable que l’excès de croissance observé soit donc simplement le reflet de cette chronobiologie normale à laquelle, toutefois, nous devrons nous opposer pour réussir le traitement des patients MH.
 |
Figure 2. Greffes de progéniteurs neuraux dérivés de CSEh chez le rat. Étude de la survie et de la maturation de greffons composés de CSEh dont la différenciation est induite pendant 45 jours in vitro, 3 à 5 mois après transplantation dans des rats modèles de la MH. (A et B) Détection de larges zones constituées de neurones humains (HNA+, en rouge) exprimant la protéine DARPP32 (en vert). (C et D) Marquage au crésyl violet d’une coupe coronale d’un cerveau de rat 3 mois après greffe (C) et immunohistochimie du marqueur de cellules neurales Pax6 et du marqueur de prolifération Ki67 (D) illustrant un greffon très volumineux constitué de cellules neurales prolifératives persistantes, 3 à 5 mois après transplantation. Barres d’échelle : 100 µm. |
En conclusion, notre étude démontre qu’il est possible, à partir de cellules souches embryonnaires humaines, de produire en laboratoire et en quantité pertinente pour une utilisation thérapeutique, des populations de cellules neurales enrichies en progéniteurs capables de se différencier en neurones striataux moyen-épineux aussi bien in vitro qu’in vivo. Cela donne une nouvelle arme à la thérapie cellulaire de la MH, susceptible de surmonter les obstacles logistiques qui en bloquent aujourd’hui l’extension. Cependant, la maîtrise de la capacité proliférative des cellules transplantées reste insuffisante, appelant à la mise en place de mesures de contrôle parfaitement efficaces avant d’envisager toute application clinique.
Remerciements
Ce travail a été soutenu par : MediCenParis Region (programme IngeCell), FP6 de l’EC (STEM-HD). Le Genopole, l’Association Huntington France et la Fédération Huntington Espoir. L.A est financée par la Fondation pour la Recherche Médicale (Prix Pomaret Delalande 2006)
Références
- Bachoud-Levi AC, Remy P, Nguyen JP, et al. Motor and cognitive improvements in patients with Huntington’s disease after neural transplantation. Lancet 2000; 356 : 1975–9. [Google Scholar]
- Bachoud-Levi AC, Gaura V, Brugieres P, et al. Effect of fetal neural transplants in patients with Huntington’s disease 6 years after surgery: a long-term follow-up study. Lancet Neurol 2006; 5 : 303–9. [Google Scholar]
- Thomson JA, Itskovitz-Eldor J, Shapiro SS, et al. Embryonic stem cell lines derived from human blastocysts. Science 1998; 282 : 1145–7. [Google Scholar]
- Aubry L, Bugi A, Lefort N, et al. Striatal progenitors derived from human ES cells mature into DARPP32 neurons in vitro and in quinolinic acid-lesioned rats. Proc Natl Acad Sci USA 2008; 105 : 16707–12. [Google Scholar]
- Perrier AL, Tabar V, Barberi T, et al. Derivation of midbrain dopamine neurons from human embryonic stem cells. Proc Natl Acad Sci USA 2004; 101 :12543–8. [Google Scholar]
- Ericson J, Muhr J, Placzek M, et al. Sonic hedgehog induces the differentiation of ventral forebrain neurons: a common signal for ventral patterning within the neural tube. Cell 1995; 81 : 747–56. [Google Scholar]
- Backman MM, Machon O, Mygland L, et al. Effects of canonical Wnt signaling on dorso-ventral specification of the mouse telencephalon. Dev Biol 2005; 279 : 155–68. [Google Scholar]
- Watanabe K, Kamiya D, Nishiyama T, et al. Directed differentiation of telencephalic precursors from embryonic stem cells. Nat Neurosci 2005; 8 : 288–96. [Google Scholar]
- Fluckiger AC, Dehay C, Savatier P. Cellules souches embryonnaires et thérapies cellulaires du système nerveux. Med Sci (Paris) 2003; 19 : 699–708. [Google Scholar]
© 2009 médecine/sciences - Inserm / SRMS
Liste des figures
|
Figure 1. Différenciation des cellules souches embryonnaires humaines en neurones striataux. A. Photo en microscopie à contraste de phase d’une colonie de cellules neurales organisées en rosettes après 23 jours de culture sur une monocouche de cellules stromales MS5. B. Après environ 62 jours de différenciation, près de 53% des neurones MAP2+ (en vert) expriment la protéine striatale DARPP32 (en rouge). Barres d’échelle : 100 µm. |
| Dans le texte | |
|
Figure 2. Greffes de progéniteurs neuraux dérivés de CSEh chez le rat. Étude de la survie et de la maturation de greffons composés de CSEh dont la différenciation est induite pendant 45 jours in vitro, 3 à 5 mois après transplantation dans des rats modèles de la MH. (A et B) Détection de larges zones constituées de neurones humains (HNA+, en rouge) exprimant la protéine DARPP32 (en vert). (C et D) Marquage au crésyl violet d’une coupe coronale d’un cerveau de rat 3 mois après greffe (C) et immunohistochimie du marqueur de cellules neurales Pax6 et du marqueur de prolifération Ki67 (D) illustrant un greffon très volumineux constitué de cellules neurales prolifératives persistantes, 3 à 5 mois après transplantation. Barres d’échelle : 100 µm. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.