")
")
| Issue |
Med Sci (Paris)
Volume 24, Number 12, Décembre 2008
|
|
|---|---|---|
| Page(s) | 1020 - 1022 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/200824121020 | |
| Published online | 15 décembre 2008 | |
Analyse dynamique de la signalisation cellulaire
L’exemple de la réponse osmotique chez Saccharomyces cerevisiae
Dynamic analysis of cell signalling: example of Saccharomyces cerevisiae osmotic answer
1
UMR CNRS 7057, Laboratoire Matière et Systèmes Complexes (MSC), Bâtiment Condorcet, Université Paris Diderot, CC 7056, 75205 Paris, France
2
FAS center for systems biology, Harvard University, Cambridge, MA, États-Unis
* Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
La cascade Hog chez la levure
Les cascades de signalisation permettent aux organismes vivants de transformer une information extracellulaire en une suite de réactions biochimiques, conduisant in fine à une réponse biologique adaptée. Les voies de signalisation de type MAPK (mitogen activated protein kinases) [1, 2] sont impliquées chez les eucaryotes dans de nombreux processus cellulaires, allant de la réponse aux stress (JNK, p38/HOG) au contrôle des processus de prolifération et de différenciation cellulaires (ERK/MAPK). Ces chemins de signalisation sont composés autour d’un bloc de trois kinases successives, formant une cascade véhiculant l’information extracellulaire depuis les récepteurs transmembranaires jusqu’au noyau, cœur de la machinerie cellulaire. En particulier, l’eucaryote modèle, Saccharomyces cerevisiae, met à profit ses voies de signalisations MAPK pour faire face à des situations mettant en péril son intégrité (choc osmotique, choc thermique) ou influençant son développement (croissance filamenteuse, sporulation) [1, 2]. Certaines de ces voies de signalisation et des protéines qui les constituent se retrouvent avec un fort degré d’homologie chez les eucaryotes supérieurs, faisant de la levure un terrain d’expériences privilégié pour l’étude des MAPK [3]. La voie de signalisation HOG (hyper osmotic glycerol) est activée en réponse à un choc hyper-osmotique et a pour homologue chez les mammifères la célèbre protéine p38. Cette voie de signalisation est très bien décrite biologiquement [3, 4] même si plusieurs interrogations persistent, d’une part sur la façon dont les récepteurs transmembranaires (Sho1p, Sln1p, Msb2p) détectent les changements de pression osmotique et, d’autre part, sur l’importance des phosphatases dans la régulation de son activité enzymatique. L’organisation structurelle de cette cascade HOG est représentée schématiquement sur la Figure 1. On y trouve un module de détection du stress osmotique - composé ici de deux unités distinctes amorcées par Sho1p [5] et Sln1p [6] - puis le bloc des trois kinases en cascade. Celui-ci est composé des MAPKKK Ste11p ou Ssk2p qui phosphoryle la MAPKK Pbs2p, celle-ci activant la phosphorylation de la MAPK Hog1p sur ses sites Thr174 et Tyr176. Après phosphorylation, cette dernière est importée dans le noyau par l’action de la karyophérine Nmd5p [7]. La translocation nucléaire de Hog1p initie une réponse très étendue au cours de laquelle près de 600 gènes sont différentiellement régulés, notamment via l’activation de plusieurs facteurs de transcription Msn2/4p, Sko1p, Hot1p, et Smp1p [8]. Les deux réponses physiologiques principales sont l’arrêt du cycle cellulaire via Sic1p et Hsl1p [9, 10] et la transcription de GPD1 et GPP2 via Hot1p [3, 4]. Ceux-ci codent pour des enzymes participant à la dégradation du glucose en glycérol, ce qui a pour effet d’augmenter la pression osmotique interne et donc de forcer la cellule à retrouver une taille compatible avec son bon fonctionnement. Après l’activation de la cascade, la concentration de hog1p dans le noyau augmente fortement, et ce n’est qu’avec le retour à l’équilibre, que, petit à petit, la protéine hog1p est déphosphorylée par les phosphatases Ptcs et Ptps [11] et exportée du noyau via la karyophérine Crm1p pour redevenir essentiellement cytoplasmique.
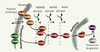 |
Figure 1. Représentation de la cascade de signalisation HOG chez Saccharomyces cerevisiae. Deux sous cascades distinctes (initiées par les protéines Sho1p et Sln1p) permettent d’activer successivement les MAPKKK, les MAPKK et finalement Hog1p, la MAP-kinase qui est transférée dans le noyau où elle initie la transcription d’un grand nombre de gènes. |
Analyse dynamique de la signalisation : questions et dispositif technique
La cascade HOG constitue un candidat idéal pour étudier quantitativement la dynamique d’une cascade de signalisation MAPK. Les questions sous-jacentes à une telle étude sont de savoir à quelle vitesse une cellule est capable de déclencher sa réponse et son adaptation à un stress extérieur ou encore, de dégager les protéines qui imposent cette dynamique. Une telle information ouvrant alors la possibilité de modifier les propriétés dynamiques d’une cascade par des constructions génétiques bien choisies.
Plutôt que d’analyser la réponse temporelle à un choc hyper osmotique bien localisé dans le temps, il est bien plus riche - mais techniquement plus difficile - de s’intéresser au comportement de cellules individuelles dans des conditions fluctuantes où le milieu nutritionnel passe périodiquement d’un état isotonique à un état hyper-osmotique, et ce pour une large gamme de fréquence. Cela revient à mesurer la réponse fréquentielle d’un système biologique. Une méthode simple et efficace consiste à utiliser une cellule de micro-fluidique (Figure 2) où deux fluides se rejoignent dans un canal de petites dimensions - quelques centaines de micromètres de largeur. À cette échelle, les deux fluides ne se mélangent pas, mais s’écoulent côte à côte formant une ligne de séparation bien définie et dont la position latérale est contrôlable en jouant sur la différence de pression entre les deux liquides (Figure 2). Avec un tel dispositif, l’environnement de cellules de levure attachées au fond de la chambre d’écoulement peut être changé rapidement et proprement [12]. Nous avons ainsi pu étudier la réponse fréquentielle de cellules de levure soumises à des chocs hyper-osmotiques périodiques, depuis des périodes très rapides (toutes les secondes) jusqu’à des périodes plus lentes (toutes les 20 minutes). Cette analyse permet d’étudier les différentes étapes de la réponse osmotique chez S. cerevisiae [12] : la réponse mécanique (variation de la taille à cause de l’augmentation de la pression osmotique externe), la réponse signalétique (localisation de la protéine Hog1p : dans le noyau) et le début de la réponse transcriptionelle (traduction de la protéine Gpd1p).
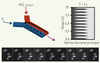 |
Figure 2. Méthode de contrôle temporel de l’environnement osmotique des cellules de levures. En jouant sur la pression d’une des deux entrées d’un canal de microfluidique, on modifie la position de l’interface des deux fluides en écoulement. Cela revient, pour des cellules situées au centre du canal, à un changement périodique de l’environnement. En utilisant un marqueur fluorescent, il est possible de suivre la position de l’interface comme le montre le diagramme spatio-temporel de droite. La séquence d’images montre le changement de localisation de la protéine Hog1p suivie par microscopie de fluorescence au cours du temps dans un environnement oscillant (T = 800 s, une image toutes les 180 s, le rayon des cellules est typiquement de quelques micromètres). |
Réponse mécanique et porosité cellulaire
À haute fréquence, le retour aux conditions standard est si rapide que même la taille des cellules ne varie pas, le transport de l’eau depuis le cytoplasme vers le milieu extracellulaire n’ayant pas le temps de se faire. Les cellules répondent de mieux en mieux aux variations à mesure que décroît la fréquence à laquelle changent les conditions environnementales : leur taille varie alors périodiquement, les cellules gonflant et se dégonflant alternativement. Faiblement d’abord puis, au-delà d’une fréquence critique, au maximum de leur capacité - qui est limitée par l’élasticité de la paroi cellulaire. Cette description correspond à celle d’un filtre passe-bas : seules les oscillations plus lentes qu’une fréquence caractéristique conduisent à une réponse, les autres, trop rapides, sont moyennées par le système, ici constitué par l’enveloppe cellulaire.
Réponse signalétique : localisation de Hog1p
Il est intéressant de noter qu’il y a découplage entre la réponse mécanique et la réponse signalétique. En effet, même si la taille des cellules suit correctement les variations environnementales pour des périodes supérieures à 10 secondes, ce n’est pas le cas de la signalisation : la protéine Hog1p, marquée par fluorescence, ne se localise pas dans le noyau, mais reste diffuse dans le cytoplasme. Ce n’est que pour des périodes supérieures à 3 minutes que l’on observe de manière reproductible la localisation et la délocalisation répétées de Hog1p dans le noyau, suivant fidèlement les fluctuations de l’environnement. La réponse transcriptionelle suit également au-delà d’une certaine période - de l’ordre de la vingtaine de minutes - les fluctuations de l’environnement et l’on observe un accroissement par paliers de l’abondance de Gpd1p, correspondant à l’accumulation de cette protéine, choc après choc.
Ingénierie de la réponse signalétique
Notre approche permet ainsi d’accéder à la dynamique des différentes observables1 de la réponse osmotique de S. cerevisiae. En particulier, il est possible de mesurer une fréquence caractéristique pour chacune de ces observables. Pour l’étape de la signalisation, cette fréquence correspond à la quantité maximale d’information qui peut transiter par unité de temps à travers la cascade MAPK depuis l’activation des récepteurs transmembranaires jusqu’au noyau. Tout comme en ingénierie, il est alors intéressant de se demander s’il est possible de changer la bande passante de ce filtre, c’est-à-dire d’accélérer ou de ralentir la signalisation cellulaire et ainsi de rendre la cellule plus ou moins sensible aux variations temporelles de son environnement. À partir de cette idée, nous avons montré qu’il était possible de ralentir considérablement la cascade de signalisation en diminuant très fortement la concentration de Pbs2p par dilution. Une étude récente s’est appuyée sur un principe expérimental similaire pour analyser la nature des boucles de rétroactions qui permettent de désactiver la cascade HOG [13]. En parallèle, nous avons montré, à l’aide de modifications génétiques bien choisies, que les deux voies d’activation de Pbs2p n’étaient pas équivalentes : la voie menée par Sln1p (phosphorelais) est ainsi plus rapide que la voie menée par l’activation de Sho1p (cascade enzymatique). Ce résultat fait écho à leurs différences architecturales, soulignant ainsi le lien entre architecture et dynamique des voies de signalisation.
Conséquences et perspectives
Notre approche, qui a vocation à s’appliquer à d’autres voies de signalisation, démontre qu’il est possible de disséquer une cascade pour d’une part, mesurer in vivo le temps nécessaire à son activation - ce qui est également une mesure de la plus lente cinétique de réaction des étapes de la cascade - et d’autre part, pour estimer l’impact de modifications génétiques sur la cinétique de cette cascade. Il serait intéressant d’observer dans quelle mesure jouer sur la concentration de ces protéines permet le contrôle de la vitesse de cette cascade. Il en ressortirait une vision dynamique de l’architecture de la cascade qui se superposerait à la vision fonctionnelle déjà existante. C’est cette vision double qui permettra d’améliorer notre compréhension de la façon dont les cascades de signalisation fonctionnent tout en développant les outils génétiques permettant d’assurer le contrôle de leur dynamique.
Remerciements
Ces recherches sont rendues possibles grâce au soutien du programme Jeunes Chercheurs de l’Agence Nationale de la recherche (PH), du « Graduate Research Program for Women Fellowship, Lucent Technologies (MNM), du « Human Frontier Science Program » (SR) et celui du FAS Center for Systems Biology (PH, SR et MNM).
Une observable est une quantité physique mesurable.
Références
- Gustin MC, Albertyn J, Alexander M, Davenport K. MAP kinase pathways in the yeast Saccharomyces cerevisiae. Microbiol Mol Biol Rev 1998; 62 : 1264–1300. [Google Scholar]
- Westfall PJ, Ballon DR, Thorner J. When the stress of your environment makes you go HOG wild. Science 2004; 306 : 1511. [Google Scholar]
- Widmann C, Gibson S, Jarpe MB, Johnson GL. Mitogen-activated protein kinase: conservation of a three-kinase module from yeast to human. Physiol Rev 1999; 79 : 143–80. [Google Scholar]
- Hohmann S. Osmotic stress signaling and osmoadaptation in yeasts. Microbiol Mol Rev 2002; 66 : 300. [Google Scholar]
- Posas F. Yeast HOG1 MAP kinase cascade is regulated by a multistep phosphorelay mechanism in the SLN1-YPD1-SSK1 two-component osmosensor. Cell 1996; 86 : 865–75. [Google Scholar]
- Posas F, Saito H. Osmotic activation of the HOG MAPK pathway via Ste11p MAPKKK : scaffold role of Pbs2p MAPKK. Science 1997; 276 : 1702–5. [Google Scholar]
- Ferrigno P, Posas F, Koepp D, et al. Regulated nucleo/cytoplasmic exchange of HOG1 MAPK requires the importin beta homologs NMD5 and XPO1. EMBO J 1998; 17 : 5606–14. [Google Scholar]
- O’Rourke SM, Herskowitz I. Unique and redundant roles for HOG MAPK pathway components as revealed by whole-genome expression analysis. Mol Biol Cell 2004; 15 : 532–42. [Google Scholar]
- Escote X, Zapater M, Clotet J, Posas F. Hog1 mediates cell-cycle arrest in G1 phase by the dual targeting of Sic1. Nat Cell Biol 2004; 6 : 997–1002. [Google Scholar]
- Clotet J, Escote X, Adrover MA, et al. Phosphorylation of Hsl1 by Hog1 leads to a G2 arrest essential for cell survival at high osmolarity. EMBO J 2006; 25 : 2338–46. [Google Scholar]
- Mattison CP, Ota IM. Two protein tyrosine phosphatases, Ptp2 and Ptp3, modulate the subcellular localization of the Hog1 MAP kinase in yeast. Genes Dev 2000; 14 : 1229–35. [Google Scholar]
- Hersen P, Mc Clean M, Mahadevan L, Ramanathan S. Signal processing by the HOG MAP kinase pathway. Proc Natl Acad Sci USA 2008; 105 : 7165–70. [Google Scholar]
- Mettetal JT, Muzzey D, Gómez-Uribe C, Van Oudenaarden A. The frequency dependence of osmo-adaptation in Saccharomyces cerevisiae. Science 2008; 319 : 482–4. [Google Scholar]
© 2008 médecine/sciences - Inserm / SRMS
Liste des figures
|
Figure 1. Représentation de la cascade de signalisation HOG chez Saccharomyces cerevisiae. Deux sous cascades distinctes (initiées par les protéines Sho1p et Sln1p) permettent d’activer successivement les MAPKKK, les MAPKK et finalement Hog1p, la MAP-kinase qui est transférée dans le noyau où elle initie la transcription d’un grand nombre de gènes. |
| Dans le texte | |
|
Figure 2. Méthode de contrôle temporel de l’environnement osmotique des cellules de levures. En jouant sur la pression d’une des deux entrées d’un canal de microfluidique, on modifie la position de l’interface des deux fluides en écoulement. Cela revient, pour des cellules situées au centre du canal, à un changement périodique de l’environnement. En utilisant un marqueur fluorescent, il est possible de suivre la position de l’interface comme le montre le diagramme spatio-temporel de droite. La séquence d’images montre le changement de localisation de la protéine Hog1p suivie par microscopie de fluorescence au cours du temps dans un environnement oscillant (T = 800 s, une image toutes les 180 s, le rayon des cellules est typiquement de quelques micromètres). |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.