")
")
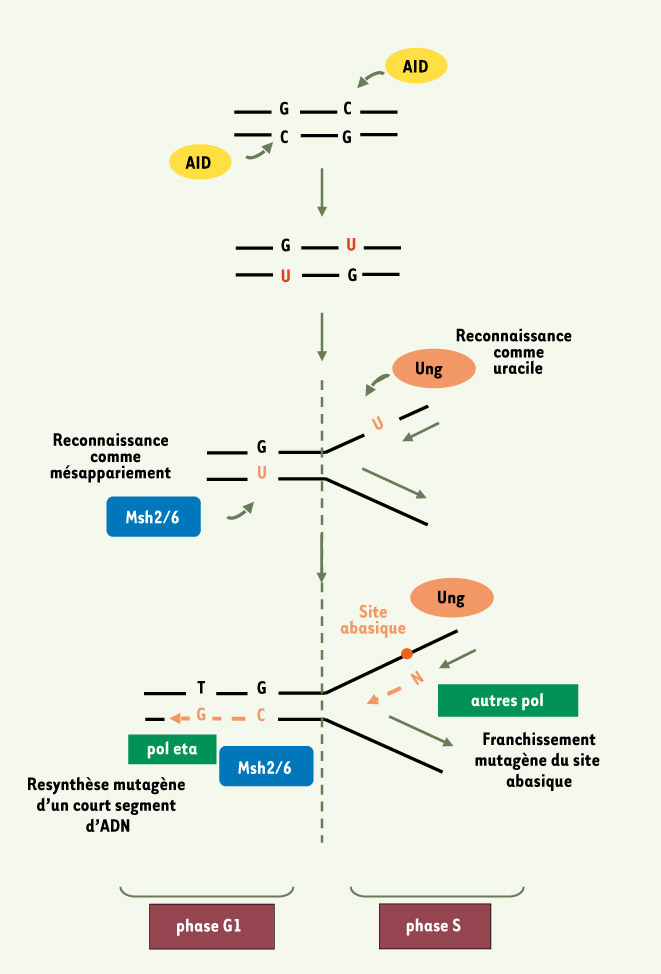
Figure 1.

Télécharger l'image originale
{kind=link}
Vue schématisée du rôle des ADN polymérases translésionnelles dans le processus d’hypermutation des gènes des immunoglobulines. L’action de l’AID au locus des immunoglobulines désamine des cytosines en uraciles. Ces uraciles présents dans l’ADN double-brin vont être préférentiellement reconnus comme des mésappariements par le complexe MSH2-MSH6 (normalement en charge de la reconnaissance des mésappariements post-réplicatifs), et empêcher ainsi le fonctionnement de la voie classique de réparation par l’uracile glycosylase UNG. Cette reconnaissance enclenche une réparation à fort taux d’erreur, par le recrutement de l’ADN polymérase eta. La signature moléculaire de cette enzyme est caractérisée par une mutagenèse sur les bases A/T, son taux d’erreurs de synthèse étant le plus fort en face d’une thymidine. Lors de la phase S, UNG pourra accéder aux uraciles exposés sur l’ADN en cours de réplication et produire des sites abasiques. La réplication de ces sites fait appel à d’autres polymérases translésionnelles (Rev1 et probablement d’autres comme pol zeta [10, 11]), dont l’action mutagène par incorporation d’une base quelconque (« N ») sera ciblée sur les bases G/C. Si le recrutement en phase S des polymérases translésionnelles correspond bien à leur rôle physiologique de franchissement des lésions bloquant la progression de la fourche de réplication, l’intervention du mismatch repair couplé à l’ADN polymérase eta apparaît, elle, comme un processus unique aux lymphocytes B engagés dans la réponse immunitaire (adapté de [8]).
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.