")
")
| Numéro |
Med Sci (Paris)
Volume 23, Numéro 5, Mai 2007
|
|
|---|---|---|
| Page(s) | 468 - 469 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/2007235468 | |
| Publié en ligne | 15 mai 2007 | |
Sur l’évolution du plan d’organisation des chordés…
On the evolution of the chordates bodyplan…
Laboratoire Arago, UMR 7628, CNRS et Université Pierre et Marie Curie, BP 44, F-66651 Banyuls sur Mer, France
L’homme partage avec tous les vertébrés, et plus largement avec tous les chordés, un certain nombre de caractéristiques morphologiques qui sont propres à leur groupe au sein du règne animal. Les plus importantes de ces caractéristiques, c’est-à-dire celles qui définissent le phylum, sont la présence d’une notochorde dorsale (d’où le nom de chordés), d’un tube neural dorsal et creux, de fentes branchiales et d’une queue post-anale. En dehors des vertébrés, le phylum des chordés est composé de deux groupes majeurs d’invertébrés, les urochordés (par exemple les ascidies) et les céphalochordés (par exemple l’amphioxus) (Figure 1) [1].
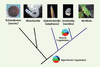 |
Figure 1. Phylogénie des deutérostomes. La branche des chordés est indiquée en bleu. Les moments clés au cours de l’évolution de l’organisateur sont indiqués par une carte schématique des territoires présomptifs. Ce sont l’apparition de l’organisateur chez l’ancêtre des chordés et la perte secondaire de l’organisateur chez les urochordés. Le bleu est utilisé pour l’ectoderme (bleu clair : épiderme ; bleu foncé : système nerveux), le vert pour le mésoderme (vert clair : mésoderme dorsal ; vert foncé : mésoderme ventral), le rouge pour l’endoderme et le noir pour l’organisateur (Figure modifiée d’après [5]). |
L’organisateur des vertébrés
L’une des questions majeures dans l’étude de l’évolution des vertébrés est de savoir quand et comment leur plan d’organisation est apparu. Pour répondre à cette question, il faut tout d’abord savoir comment ce plan d’organisation se met en place au cours de l’embryogenèse. L’une des étapes clés à cet égard a été la découverte, par Spemann en 1924 [2], d’une région chez l’embryon des vertébrés, plus particulièrement la partie dorsale du blastopore de la gastrula précoce, responsable d’organiser les axes corporels et le développement des structures morphologiques fondamentales caractéristiques des chordés. Cette région a été appelée « l’organisateur ». Si l’on enlève chirurgicalement les cellules qui composent l’organisateur d’une gastrula précoce de vertébré, aucune des caractéristiques des chordés ne se forme (tube neural dorsal et creux, notocorde, fentes branchiales). De plus, si l’on greffe ces cellules en un site différent d’une gastrula au même stade, cela induit la formation de ces structures de manière ectopique. L’organisateur contrôle le développement de l’embryon par un phénomène d’induction des tissus voisins qui détermine leur devenir cellulaire tant au niveau de leur position (antéro-postérieure et dorso-ventrale) que de leur différenciation (tissu neural, notocorde, somites, etc.). Il semble donc clair que l’apparition des vertébrés a dû être liée à l’apparition de cette région organisatrice au cours de la gastrulation chez l’un de leurs ancêtres.
Absence d’organisateur chez les urochordés
Mais comment les chordés non vertébrés établissent-ils leur plan d’organisation ? Historiquement, les urochordés ont toujours été placés à la base des chordés et leur développement embryonnaire a longtemps été considéré comme « déterminatif », tandis que celui des vertébrés, comme nous l’avons montré plus haut, est inductif. En effet, certaines expériences de manipulation de blastomères isolés d’ascidie ont permis aux chercheurs d’affirmer que le phénomène d’induction était beaucoup moins important chez les urochordés que chez les vertébrés. Cependant, des interactions inductives existent au cours de l’embryogenèse des ascidies (par exemple, spécification de l’endoderme par des signaux dorso-ventraux, induction de la notocorde par l’endoderme ou induction neurale par la corde neurale) [3]. Néanmoins, il est très clair que les urochordés ne possèdent pas de structure ayant les caractéristiques fonctionnelles de l’organisateur des vertébrés. En effet, si l’on analyse chez les ascidies le patron d’expression des gènes qui s’expriment au niveau de l’organisateur des vertébrés (par exemple Chordin, ADMP, Nodal, Lefty, Goosecoid et Lim1/5), on n’observe pas de conservation [4]. Cette absence d’organisateur est-elle un caractère dérivé chez les ascidies ou bien est-elle le caractère ancestral des chordés ? Jusqu’à récemment, cette question restait encore sans réponse.
Phylogénie des chordés et caractère ancestral de l’organisateur
La position phylogénétique relative des deux groupes d’invertébrés chordés (urochordés et céphalochordés) par rapport aux vertébrés a récemment été revisitée. L’accumulation de séquences dans les bases de données obtenues grâce aux projets de séquençage complet des génomes d’ascidie et d’amphioxus a permis la réalisation d’études phylogénétiques poussées qui placent aujourd’hui les céphalochordés, auparavant considérés comme groupe frère des vertébrés, à la base de la lignée évolutive des chordés (Figure 1) [5]. Ce changement de notre vision de la phylogénie des chordés est très important dans notre compréhension du caractère ancestral ou non des différents aspects du développement embryonnaire chez les chordés. En effet, la comparaison entre l’amphioxus et les vertébrés peut nous permettre de savoir si un mécanisme développemental est commun à tous les chordés ou s’il est spécifique des vertébrés. C’est ainsi que très récemment le groupe de Linda Holland a entrepris l’étude de l’expression de différents gènes orthologues des gènes marqueurs de l’organisateur des vertébrés chez l’embryon d’amphioxus [6]. Ces études montrent que l’expression au cours du développement de la plupart de ces gènes est comparable chez l’amphioxus à ce que l’on observe chez les vertébrés. De plus, un traitement exogène des embryons avec la protéine BMP4 (normalement localisée ventralement chez la gastrula des vertébrés, et possédant une fonction antineurale) entraîne une ventralisation de l’embryon d’amphioxus, ce qui démontre que du point de vue fonctionnel, l’établissement de l’axe dorso-ventral (D/V) est aussi conservé entre l’amphioxus et les vertébrés [6]. Ces résultats confortent d’autres résultats antérieurs obtenus par Tung et ses collaborateurs dans les années 1960 [7], qui démontrèrent par des expériences de transplantation de blastomères les capacités inductives de la partie dorsale du blastopore de la gastrula d’amphioxus. Ainsi, il semble désormais clair que l’ancêtre de tous les chordés établissait son plan d’organisation par l’intermédiaire de la fonction inductrice d’un organisateur au cours de la gastrulation et que cette fonction organisatrice a ensuite été perdue chez les ascidies. D’autres données laissent également penser qu’au moins une partie des mécanismes d’organisation devait déjà être présente avant l’apparition des chordés. En effet, chez les hémichordés, l’un des phylums le plus proche des chordés et qui partage avec eux la présence de structures typiquement induites par l’organisateur telles que les fentes branchiales, le mésoderme du pharynx ou l’endostyle, on observe déjà une expression dorso-ventrale de BMP et de ses antagonistes (par exemple, la chordine) mais de manière inversée par rapport aux chordés [8]. Chez les protostomiens, tels que la drosophile, les orthologues de BMP/Chordine sont aussi exprimés dans l’axe D/V des embryons avec une orientation similaire à celle qui prévaut chez les hémichordés [9]. Ces résultats suggèrent que le plan d’organisation typique des vertébrés soit apparu spécifiquement chez l’ancêtre de tous les chordés, après l’inversion de l’axe D/V, inversion qui doit être étroitement liée à l’évolution fonctionnelle de l’organisateur.
Aujourd’hui, deux questions restent donc sans réponse. La première concerne les mécanismes moléculaires qui ont dû être modifiés pour permettre l’apparition fonctionnelle de l’organisateur à partir d’un ancêtre ayant un plan d’organisation possédant l’axe D/V inversé par rapport à celui des vertébrés. La deuxième est de savoir quelles sont les adaptations chez les ascidies qui leur ont permis de garder un plan d’organisation de type chordé en l’absence d’un véritable organisateur.
Références
- Schubert M, Escriva H, Xavier-Neto J, Laudet V. Amphioxus and tunicates as evolutionary model systems. Trends Ecol Evol 2006; 21 : 269–77. [Google Scholar]
- Spemann H, Mangold H. Über Induktion von Embryonalanlagen durch Implantation artfremder Organisatoren. Dev Genes Evol 1924; 100 : 599–638. [Google Scholar]
- Kourakis MJ, Smith WC. Did the first chordates organize without the organizer ? Trends Genet 2005; 21 : 506–10. [Google Scholar]
- Imai KS, Hino K, Yagi K, et al. Gene expression profiles of transcription factors and signaling molecules in the ascidian embryo: towards a comprehensive understanding of gene networks. Development 2004; 131 : 4047–58. [Google Scholar]
- Delsuc F, Brinkmann H, Chourrout D, Philippe H. Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature 2006; 439 : 965–8. [Google Scholar]
- Yu JK, Satou Y, Holland ND, et al. Axial patterning in cephalochordates and the evolution of the organizer. Nature 2007; 445 : 613–7. [Google Scholar]
- Tung TC, Wu SC, Tung YYF. Experimental studies on the neural induction in Amphioxus. Scientia Sinica 1962; XI: 805–20. [Google Scholar]
- Lowe CJ, Terasaki M, Wu M, et al. Dorsoventral patterning in hemichordates: insights into early chordate evolution. PLoS Biol 2006; 4 : e291. [Google Scholar]
- Dorfman R, Shilo BZ. Biphasic activation of the BMP pathway patterns the Drosophila embryonic dorsal region. Development 2001; 128 : 965–72. [Google Scholar]
© 2007 médecine/sciences - Inserm / SRMS
Liste des figures
|
Figure 1. Phylogénie des deutérostomes. La branche des chordés est indiquée en bleu. Les moments clés au cours de l’évolution de l’organisateur sont indiqués par une carte schématique des territoires présomptifs. Ce sont l’apparition de l’organisateur chez l’ancêtre des chordés et la perte secondaire de l’organisateur chez les urochordés. Le bleu est utilisé pour l’ectoderme (bleu clair : épiderme ; bleu foncé : système nerveux), le vert pour le mésoderme (vert clair : mésoderme dorsal ; vert foncé : mésoderme ventral), le rouge pour l’endoderme et le noir pour l’organisateur (Figure modifiée d’après [5]). |
| Dans le texte | |
Les statistiques affichées correspondent au cumul d'une part des vues des résumés de l'article et d'autre part des vues et téléchargements de l'article plein-texte (PDF, Full-HTML, ePub... selon les formats disponibles) sur la platefome Vision4Press.
Les statistiques sont disponibles avec un délai de 48 à 96 heures et sont mises à jour quotidiennement en semaine.
Le chargement des statistiques peut être long.