")
")
| Issue |
Med Sci (Paris)
Volume 21, Number 5, Mai 2005
|
|
|---|---|---|
| Page(s) | 467 - 469 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/2005215467 | |
| Published online | 15 mai 2005 | |
Un nouveau partenaire de MuSK à la junction neuromusculaire, la 14-3-3 γ, réprime la transcription des génes synaptiques
14-3-3 γ, a novel partner of MuSK downregulates synaptic gene transcription at the neuromuscular junction
Biologie cellulaire des membranes, Institut Jacques Monod, CNRS UMR 7592, Universites Paris 6 et Paris 7, 2, place Jussieu, 75251 Paris Cedex 05, France
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Les jonctions neuromusculaires s’établissent à la suite d’interactions complexes entre les terminaisons axonales des motoneurones, les fibres musculaires et les cellules de Schwann. La genèse de la jonction neuromusculaire culmine avec la formation de deux domaines spécialisés du nerf et du muscle, la terminaison présynaptique impliquée dans la libération de l’acétylcholine (ACh), et l’appareil postsynaptique responsable de la reconnaissance de l’ACh et de la dépolarisation de la membrane. La membrane postsynaptique assure ces fonctions sur la base d’une accumulation de récepteurs de l’ACh. L’efficacité de la transmission synaptique est renforcée par la juxtaposition précise des sites de libération de l’ACh (les zones actives présynaptiques) et des récepteurs postsynaptiques qui assure leur activation rapide (de l’ordre de la microseconde). Une telle organisation suggère que des mécanismes complexes de signalisation entre le motoneurone et la fibre musculaire sont nécessaires à la coordination de la différenciation pré- et postsynaptique.
La formation de l’appareil postsynaptique requiert, outre le récepteur de l’ACh, au minimum trois composants : l’agrine, un héparane sulfate protéoglycane sécrété par le nerf, un récepteur tyrosine kinase spécifique du muscle (MuSK), la pièce centrale du complexe récepteur de l’agrine, et la rapsyne, une protéine extrinsèque nécessaire à l’agrégation du récepteur de l’ACh [1]. L’activation de MuSK (par phosphorylation de tyrosines) par l’agrine déclenche en aval une cascade d’événements aboutissant au recrutement du récepteur de l’ACh via la rapsyne. L’invalidation chez la souris des gènes codant pour chacun de ces trois composants entraîne des perturbations majeures de la synaptogenèse, notamment l’absence de différenciation postsynaptique [2]. La jonction neuromusculaire comprend aussi une spécialisation du cytosquelette et des noyaux sous-neuraux encore appelés noyaux fondamentaux. Certains gènes musculaires tels que les gènes codant pour les sous-unités du récepteur de l’ACh sont transcrits préférentiellement par ces noyaux sous-neuraux, entraînant une concentration locale des messagers correspondants [3]. Deux facteurs exprimés dans les motoneurones et libérés dans l’espace synaptique, la neuréguline (NRG)-1 et l’agrine sont susceptibles d’être impliqués dans ce processus. Dans les myotubes en culture, l’activation des récepteurs ErbB (RTK de la famille du récepteur de l’epidermal growth factor) par la NRG stimule la transcription des gènes des sous-unités α, δet ε du récepteur de l’Ach et celle d’autres gènes codant pour des protéines synaptiques (utrophine, rapsyne…) via les voies de la MAP-kinase (ERK), c-Jun kinase (JNK) et de la phosphatidyl inositol-3 kinase (PI3K). Ces voies phosphorylent en aval le facteur de transcription GA-binding protein (GABP) qui se fixe sur une séquence régulatrice spécifique des gènes synaptiques, la boîte N. Cependant, le rôle de la NRG neuronale dans l’activation des gènes codant pour le récepteur de l’ACh reste à démontrer in vivo. En revanche, l’activation MuSK in vivo est suffisante pour induire la formation d’un appareil postsynaptique ectopique incluant la transcription compartimentée des gènes codant pour les récepteurs de l’ACh en l’absence de terminaison nerveuse, donc de NRG neuronale [4].
La recherche des partenaires de MuSK impliqués dans l’agrégation des protéines synaptiques, ou dans la transcription de ces gènes, est une étape indispensable pour comprendre les mécanismes de la différenciation de la jonction neuromusculaire. Les recherches menées par différentes équipes ont conduit à la découverte de l’adaptateur moléculaire dishevelled (Dvl1) [5], d’une molécule de signalisation, la tyrosine kinase Abelson [6], et de la géranylgéranyltransférase I [7]. Ces différentes molécules jouent un rôle clé dans le recrutement du récepteur de l’ACh en participant à la réorganisation du réseau d’actine cortical via les petites GTPases de la famille Rho (Rac/Cdc 42) et la sérine/thréonine kinase PAK1. Ces GTPases, requises dans le mécanisme d’agrégation du récepteur de l’ACh [6], activent également la transcription synaptique en stimulant la voie JNK en aval du complexe Agrine/MuSK [9].
Au sein de notre équipe, nous avons recherché les différents composants du complexe MuSK par une approche protéomique fondée sur le pontage chimique de MuSK à ses partenaires dans son environnement membranaire (membrane postsynaptique purifiée à partir des organes électriques du poisson torpille). Les complexes de MuSK solubilisés ont ensuite été purifiés par immunoaffinité. Après séparation, les produits de pontage de MuSK ont été analysés par spectrométrie de masse MALDI-TOF. Parmi plusieurs partenaires potentiels de MuSK, nous avons identifié la protéine 14-3-3 γ [10]. Les protéines 14-3-3 constituent une famille émergente d’adaptateurs moléculaires impliqués dans de nombreux processus de signalisation cellulaire [11]. Elles fonctionnent en interagissant avec de nombreuses cibles moléculaires (plus de 100 sont aujourd’hui connues). La protéine 14-3-3 γ, seule isoforme exprimée dans le muscle, est co-localisée avec les récepteurs de l’ACh à la jonction neuromusculaire. En collaboration avec les équipes de Laurent Schaeffer (ENS, Lyon, France) et de Jean-Pierre Changeux (Institut Pasteur, Paris, France), nous avons montré que la surexpression de la 14-3-3 γ dans des myotubes en culture ou dans les fibres musculaires in vivo réprime spécifiquement l’expression de plusieurs gènes synaptiques [10]. De plus, après électroporation in vivo de 1a 14-3-3 γ dans les muscles de souris, la distribution des agrégats de récepteur de l’ACh dans la membrane postsynaptique est anormale. Parmi les cibles connues des protéines 14-3-3, se trouvent deux effecteurs de la voie NRG/ErbB, la PI3K et Raf-1, une kinase de la voie ERK. Nous avons mis en évidence le fait que la surexpression de la 14-3-3 γ inhibe la transcription des gènes rapporteurs synaptiques, induite par PI3K ou Raf-1 constitutivement actifs, dans les myotubes en culture [10]. L’ensemble de ces résultats indique que le complexe MuSK/14-3-3 γ régule l’expression des gènes synaptiques à la jonction neuromusculaire (Figure 1). Cette découverte renforce l’hypothèse selon laquelle MuSK est l’élément central dans la formation et le maintien de la synapse, responsable à la fois de la localisation des protéines de la membrane postsynaptique et de la régulation de l’expression des gènes synaptiques. Dans cette hypothèse, la 14-3-3 γ serait le lien entre MuSK et la cascade de signalisation NRG/ErbB, cela suggérant la possibilité d’un dialogue entre les voies activées par la NRG et l’agrine (Figure 2). Restent à découvrir les mécanismes de régulation de l’interaction MuSK/14-3-3 γ. Le domaine cytoplasmique de MuSK, comme beaucoup de ligands des protéines 14-3-3, possède deux séquences consensus de liaison (RSxSxP et Rx1-2Sx2-3S) qui contiennent des résidus sérine. La phosphorylation d’au moins un de ces résidus est nécessaire à la liaison de la 14-3-3 à ses ligands. On peut donc imaginer que des signaux différents de l’agrine contrôlent l’interaction MuSK-14-3-3 γ
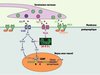 |
Figure 1. Le complexe MuSK/14-3-3 γà la jonction neuromusculaire. La protéine 14-3-3 γ inhibe la transcription des gènes synaptiques en interférant avec la voie de signalisation NRG/ErbB, en bloquant la protéine Raf-1 dans une conformation inactive. Cette inhibition empêche l’activation de la voie ERK, la phosphorylation de la GA-binding protein (GABP) et sa fixation sur la boîte N (N) des promoteurs des gènes synaptiques. La 14-3-3 γ étant active sous forme dimérique, les sites respectifs d’interaction avec MuSK ou Raf-1 sont probablement portés par chaque monomère. La voie de la phosphatidyl inositol-3 phosphate kinase n’est pas représentée. Ach : acétylcholine ; E : boîte E, site de fixation des facteurs de transcription myogéniques (MyoD…) ; RACh : récepteur de l’acétylcholine. |
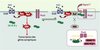 |
Figure 2. Dialogue entre les voies de signalisation de MuSK et ErbB. Le recrutement de la 14-3-3 γ par MuSK positionnerait cette dernière près de la membrane où elle serait susceptible d’inactiver Raf-1, un effecteur de la voie ERK stimulé par NRG/ErbB. La phosphorylation de sérines présentes dans les sites consensus de liaison de la 14-3-3 du domaine cytoplasmique de MuSK, serait induite par un signal encore non identifié, mais différent de l’agrine. Les astérisques blanches dans le domaine cytoplasmique de MuSK indiquent les sérines potentiellement phosphorylées. Dans le cartouche sont détaillées les deux séquences consensus de liaison de la 14-3-3 γ du domaine cytoplasmique de MuSK. |
Références
- Ruegg MA, Bixby JL. Agrin orchestrates synaptic differentiation at the vertebrate neuromuscular junction. Trends Neurosci 1998; 21 : 22–7. [Google Scholar]
- Sanes J, Litchman JW. Induction, assembly, maturation and maintenance of a postsynaptic apparatus. Nat Rev Neurosci 2001; 2 : 791–805. [Google Scholar]
- Schaeffer L, de Kerchove d’Exaerde A, Changeux JP. Targeting transcription to the neuromuscular synapse. Neuron 2001; 31 : 15–22. [Google Scholar]
- Jones G, Moore C, Hashemolhosseini S, Brenner HR. Constitutively active MuSK is clustered in the absence of agrin and induces ectopic postsynaptic-like membranes in skeletal muscle fibers. J Neurosci 1997; 19 : 3376–83. [Google Scholar]
- Luo ZG, Wang Q, Zhou JZ, et al. Regulation of AChR clustering by Dishevelled interacting with MuSK and PAK 1. Neuron 2002; 35 : 489–505. [Google Scholar]
- Finn AJ, Feng G, Pendergast AM. Postsynaptic requirement for Abl kinases in assembly of the neuromuscular junction. Nat Neurosci 2003; 7 : 717–23. [Google Scholar]
- Luo ZG, Je HS, Wang Q, et al. Implication of geranylgeranyltransferase I in synapse formation. Neuron 2003; 40 : 703–17. [Google Scholar]
- Weston C, Gordon C, Teressa G, et al. Cooperative regulation by Rac and Rho of agrin-induced acetylcholine receptor clustering in muscle cells. J Biol Chem 2003; 278 : 6450–5. [Google Scholar]
- Lacazette E, Le Calvez S, Gajendran N, Brenner HR. A novel pathway for MuSK to induce key genes in neuromuscular synapse formation. J Cell Biol 2003; 161 : 727–36. [Google Scholar]
- Strochlic L, Cartaud A, Mejat A, et al. 14-3-3 γ associates with muscle-specific kinase and regulates synaptic gene transcription at vertebrate neuromuscular synapse. Proc Natl Acad Sci USA 2004; 101 : 18189–94. [Google Scholar]
- Van Hemert MJ, de Steensma HY, van Heusden GPH. 14-3-3 proteins: key regulators of cell division, signalling and apoptosis. BioEssays 2001; 23 : 936–46. [Google Scholar]
© 2005 médecine/sciences - Inserm / SRMS
Liste des figures
|
Figure 1. Le complexe MuSK/14-3-3 γà la jonction neuromusculaire. La protéine 14-3-3 γ inhibe la transcription des gènes synaptiques en interférant avec la voie de signalisation NRG/ErbB, en bloquant la protéine Raf-1 dans une conformation inactive. Cette inhibition empêche l’activation de la voie ERK, la phosphorylation de la GA-binding protein (GABP) et sa fixation sur la boîte N (N) des promoteurs des gènes synaptiques. La 14-3-3 γ étant active sous forme dimérique, les sites respectifs d’interaction avec MuSK ou Raf-1 sont probablement portés par chaque monomère. La voie de la phosphatidyl inositol-3 phosphate kinase n’est pas représentée. Ach : acétylcholine ; E : boîte E, site de fixation des facteurs de transcription myogéniques (MyoD…) ; RACh : récepteur de l’acétylcholine. |
| Dans le texte | |
|
Figure 2. Dialogue entre les voies de signalisation de MuSK et ErbB. Le recrutement de la 14-3-3 γ par MuSK positionnerait cette dernière près de la membrane où elle serait susceptible d’inactiver Raf-1, un effecteur de la voie ERK stimulé par NRG/ErbB. La phosphorylation de sérines présentes dans les sites consensus de liaison de la 14-3-3 du domaine cytoplasmique de MuSK, serait induite par un signal encore non identifié, mais différent de l’agrine. Les astérisques blanches dans le domaine cytoplasmique de MuSK indiquent les sérines potentiellement phosphorylées. Dans le cartouche sont détaillées les deux séquences consensus de liaison de la 14-3-3 γ du domaine cytoplasmique de MuSK. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.