")
")
| Issue |
Med Sci (Paris)
Volume 21, Number 5, Mai 2005
|
|
|---|---|---|
| Page(s) | 465 - 467 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/2005215465 | |
| Published online | 15 mai 2005 | |
Un nouveau modèle de production des cellules souches hématopoïétiques intra-embryonnaires
A new model for intra-embryonic hematopoietic stem cell generation
1
Unite de Developpement des lymphocytes, Inserm U.668, Institut Pasteur, 25, rue du Docteur Roux, 75724 Paris Cedex 15, France
2
Inserm U.362, Institut Gustave Roussy, 39, rue Camille Desmoulins, 94805 Villejuif, France
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Développement du système hématopoïétique
Chez l’adulte, l’ensemble des cellules sanguines est constamment renouvelé à partir d’une population de cellules souches hématopoïétiques (CSH) faiblement représentées dans la moelle osseuse. Les CSH adultes sont supposées dériver d’une population produite pendant le développement embryonnaire, suivant des modalités encore mal connues.
Un ensemble de travaux menés depuis une dizaine d’années dans de nombreux modèles animaux a permis d’établir qu’il existe deux générations indépendantes de précurseurs hématopoïétiques au cours de l’ontogenèse. La première, qui a lieu au niveau du sac vitellin, extra-embryonnaire, fournit rapidement des globules rouges, des macrophages et des mégacaryocytes, mais ces précurseurs ont un potentiel de différenciation limité et sont incapables de maintenir, à long terme, la production de cellules sanguines. Les précurseurs formés un peu plus tardivement dans le compartiment intra-embryonnaire (dans une région qui comprend l’aorte, les gonades et le mésonéphros, appelée AGM) possèdent, quant à eux, les caractéristiques de multipotentialité et d’auto-renouvellement à long terme qui caractérisent les CSH [1].
Bien que les données cytologiques et fonctionnelles obtenues in vitro et in vivo s’accordent pour désigner l’aorte et le mésenchyme sous-jacent comme le lieu d’origine des CSH, la localisation précise de leur site d’émergence n’a pas encore été déterminée. L’analyse cytologique de l’AGM au moment de la production de CSH (pour revue, voir [1]), montre l’existence d’agrégats composés de cellules hématopoïétiques immatures, localisés sur les parois ventro-latérales de l’aorte, et de plages cellulaires localisées sous les îlots aortiques, qui expriment, entre autres, l’antigène AA4.1 et le facteur de transcription GATA-3, et qui n’expriment pas le marqueur caractéristique des cellules hématopoïétiques CD45. Alors que les « îlots aortiques » ont été décrits chez les embryons de tous les vertébrés, y compris l’embryon humain, les plages cellulaires ont été caractérisées jusqu’à présent uniquement chez l’embryon de souris [2] et chez l’humain [3].
Comment se forment les CSH intra-embryonnaires ?
La nature des processus conduisant à la production des CSH au niveau de l’aorte fait actuellement l’objet de controverses.
L’ensemble des données cytologiques (pour revue, voir [1]), indique que les précurseurs intra-embryonnaires, com-me ceux du sac vitellin, expriment un grand nombre de protéines et de gènes également exprimés par les cellules endothéliales (CD31, CD34, c-kit, AA4.1, GATA-2, entre autres). Par ailleurs, des expériences pratiquées in vivo et in vitro ont montré que les CSH intra-embryonnaires expriment c-kit, CD34, CD31 et AA4.1. Enfin, des données plus récentes indiquent que des cellules qui n’expriment pas CD45 sont capables de produire des cellules sanguines de tous les lignages. L’absence d’expression de CD45, jumelée à l’expression de marqueurs endothéliaux ont conduit à considérer que l’endothélium aortique pouvait donner naissance aux CSH intra-embryonnaires (modèle de « l’endothélium hématogène ») [4–8].
Apports d’une étude multiparamétrique : phénotype, fonction et expression
Nous avons caractérisé [9], par cytométrie de flux, dans l’AGM d’embryons de souris, une population de cellules CD45-, c-kit+, AA4.1+, CD31+, CD41+. Cette population comprend la quasi-totalité des précurseurs multipotents précédemment quantifiés dans l’AGM. Il s’agit bien de CSH, puisque ces cellules sont capables de reconstituer à long terme le système hématopoïétique de souris a-lymphoïdes, dépourvues de cellules natural killer.
L’analyse des CSH purifiées à partir de l’AGM montre qu’elles expriment des marqueurs de surface (CD31, CD34, c-kit, AA4.1) et des facteurs de transcription (Lmo2, GATA-2) communs aux lignages hématopoïétiques et endothéliaux. Mais elles se distinguent des cellules endothéliales par l’expression de CD41, récemment identifié comme un marqueur des précurseurs hématopoïétiques embryonnaires [10], et du facteur de transcription GATA-3, tous deux absents des cellules endothéliales.
L’ensemble des marqueurs de CSH ainsi définis a été corrélé avec les profils d’expression précédemment obtenus in situ chez l’embryon [2] et complété par une cartographie phénotypique en microscopie confocale [9]. La combinaison de marqueurs qui définissent les CSH est bien localisée au niveau des îlots aortiques, mais également par des cellules localisées au sein des plages sous-aortiques (Figure 1).
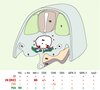 |
Figure 1. Distribution des gènes et marqueurs membranaires exprimés par les cellules hématopoïétiques et endothéliales dans la région aortique de l’embryon de souris. La combinaison de gènes et marqueurs de surfaces exprimés par les cellules souches hématopoïétiques (CSH, en orange) purifiées à partir d’une région appelée AGM, qui comprend l’aorte (Ao), les gonades (G) et le mésonéphros (M), révèle exclusivement des cellules faisant partie des îlots aortiques ou des plages sous-aortiques. Les flèches bleues indiquent le sens de migration présomptif des CSH, depuis les plages sous-aortiques (PSA, en vert) vers la lumière de l’aorte où elles contribuent aux îlots, avant d’être libérées dans la circulation et de coloniser le foie fœtal. CE : cellules endothéliales (en mauve) ; CH (MΦ) : cellules hématopoïétiques, macrophages (en rouge) ; Int : intestin ; Mn : mésentère ; S : somites ; TN : tube neural. |
Définition d’un modèle alternatif de production des CSH
L’ensemble des données que nous avons obtenues [9] montrent que les CSH, identifiées par leur phénotype (CD45-, c-kit+, AA4.1+, CD41+, CD31+) sont localisées, non seulement dans le plancher aortique, mais aussi dans le mésenchyme sub-aortique. Ces résultats désignent donc les plages sous-aortiques comme le site de production des CSH, et non pas le plancher aortique, où la présence de CSH semble plutôt refléter leur libération dans la circulation sanguine, préalable à la colonisation du foie fœtal. Les perspectives ouvertes par ce travail [9], et en particulier la caractérisation de la fonction des composants des plages sous-aortiques dans la formation des CSH, pourraient déboucher sur l’identification de signaux impliqués dans la différenciation de cellules mésodermiques en dérivés hématopoïétiques. Ces connaissances seront fondamentales pour la manipulation des cellules souches embryonnaires dans leur différenciation vers la voie hématopoïétique.
Références
- Godin I, Cumano A. The hare and the tortoise : an embryonic haematopoietic race. Nat Rev Immunol 2002; 2 : 593–604. [Google Scholar]
- Manaia A, Lemarchandel V, Klaine M, et al. Lmo2 and GATA-3 associated expression in intraembryonic hemogenic sites. Development 2000; 127 : 643–53. [Google Scholar]
- Marshall CJ, Moore RL, Thorogood P, et al. Detailed characterization of the human aorta-gonad-mesonephros region reveals morphological polarity resembling a hematopoietic stromal layer. Dev Dyn 1999; 215 : 139–47. [Google Scholar]
- Jaffredo T, Gautier R, Eichmann A, Dieterlen-Lievre F. Intraaortic hemopoietic cells are derived from endothelial cells during ontogeny. Development 1998; 125 : 4575–83. [Google Scholar]
- Nishikawa SI, Nishikawa S, Kawamoto H, et al. In vitro generation of lymphohematopoietic cells from endothelial cells purified from murine embryos. Immunity 1998; 8 : 761–9. [Google Scholar]
- De Bruijn MF, Ma X, Robin C, et al. Hematopoietic stem cells localize to the endothelial cell layer in the midgestation mouse aorta. Immunity 2002; 16 : 673–83. [Google Scholar]
- North TE, de Bruijn MF, Stacy T, et al. Runx1 expression marks long-term repopulating hematopoietic stem cells in the midgestation mouse embryo. Immunity 2002; 16 : 661–72. [Google Scholar]
- Oberlin E, Tavian M, Blazsek I, Peault B. Blood-forming potential of vascular endothelium in the human embryo. Development 2002; 129 : 4147–57. [Google Scholar]
- Bertrand JY, Giroux S, Golub R, et al. Characterization of purified intraembryonic hematopoietic stem cells as a tool to define their site of origin. Proc Natl Acad Sci USA 2005; 102 : 134–9. [Google Scholar]
- Mikkola HK, Fujiwara Y, Schlaeger TM, et al. Expression of CD41 marks the initiation of definitive hematopoiesis in the mouse embryo. Blood 2003; 101 : 508–16. [Google Scholar]
© 2005 médecine/sciences - Inserm / SRMS
Liste des figures
|
Figure 1. Distribution des gènes et marqueurs membranaires exprimés par les cellules hématopoïétiques et endothéliales dans la région aortique de l’embryon de souris. La combinaison de gènes et marqueurs de surfaces exprimés par les cellules souches hématopoïétiques (CSH, en orange) purifiées à partir d’une région appelée AGM, qui comprend l’aorte (Ao), les gonades (G) et le mésonéphros (M), révèle exclusivement des cellules faisant partie des îlots aortiques ou des plages sous-aortiques. Les flèches bleues indiquent le sens de migration présomptif des CSH, depuis les plages sous-aortiques (PSA, en vert) vers la lumière de l’aorte où elles contribuent aux îlots, avant d’être libérées dans la circulation et de coloniser le foie fœtal. CE : cellules endothéliales (en mauve) ; CH (MΦ) : cellules hématopoïétiques, macrophages (en rouge) ; Int : intestin ; Mn : mésentère ; S : somites ; TN : tube neural. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.