")
")
| Numéro |
Med Sci (Paris)
Volume 21, Numéro 1, Janvier 2005
|
|
|---|---|---|
| Page(s) | 7 - 9 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/20052117 | |
| Publié en ligne | 15 janvier 2005 | |
Le mécanisme d’action de l’hepcidine déchiffré
Deciphering the action mechanism of hepcidin
1
Inserm U.409, Faculté de Médecine Xavier Bichat, 16, rue Henri Huchard, BP 416, 75870 Paris Cedex, France
2
Inserm U.567, Institut Cochin, 24, rue du Faubourg Saint-Jacques, 75014 Paris Cedex, France
L’homéostasie du fer dans l’organisme repose sur un contrôle strict de l’absorption du fer au niveau des entérocytes matures de la villosité duodénale. Ce processus met en jeu un grand nombre de protéines dont des transporteurs de fer capables de faire traverser le métal du côté apical (la protéine DMT1, divalent metal transpoter) et du côté basolatéral de la cellule (la ferroportine) (Figure 1) (pour revue, voir [1]). L’hepcidine est une petite protéine synthétisée par l’hépatocyte sous la forme d’un précurseur de 84 acides aminés. La maturation de l’hepcidine après clivage du précurseur conduit à deux formes moléculaires de 25 et 20 acides aminés. Ces formes matures, correspondant à la partie carboxy-terminale, ont été isolées et purifiées dans le sérum et dans l’urine. Elles possèdent huit cystéines formant quatre ponts disulfures qui leur confèrent une structure compacte unique. D’abord identifiée pour son activité antimicrobienne, l’hepcidine s’est révélée être un peptide hormonal clé du métabolisme du fer capable d’inhiber l’absorption intestinale du fer alimentaire. Un régime riche en fer a pour conséquence d’augmenter la production d’hepcidine afin de diminuer l’absorption de ce fer en excès [2, 3]. Dans les situations d’hémochromatoses sévères (lorsque par exemple l’hepcidine est mutée, que ce soit chez l’homme ou chez la souris), cette régulation de l’absorption intestinale de fer est perdue, ce qui provoque une accumulation de fer très importante dans les organes [2, 4]. À l’inverse, une surexpression d’hepcidine crée un blocage de l’absorption de fer, ce qui entraîne une carence en fer pouvant aboutir à une anémie [5]. Si l’on connaît bien maintenant le rôle que joue l’hepcidine dans le contrôle de l’homéostasie du fer, on ignorait encore tout de son mode d’action pour inhiber l’absorption du fer, et surtout sur quelle cible moléculaire elle était capable d’agir. L’exporteur de fer évoqué ci-dessus, la ferroportine, protéine transmembranaire présente dans la membrane basolatérale des entérocytes, semblait être une cible rêvée. C’est effectivement ce que vient de démontrer le groupe de Jerry Kaplan (Université d’Utah, USA) en collaboration avec le groupe de Tomas Ganz (Université de Californie, USA) dans une étude publiée dans Science [6]. Les auteurs ont utilisé un modèle cellulaire (cellules humaines HEK293) dans lequel la ferroportine fusionnée à la GFP (green fluorescent protein), la ferro-GFP, est exprimée de manière inductible. En présence d’inducteur, la ferro-GFP produite se localise dans la membrane et provoque, lorsque les cellules sont incubées en présence de fer, une diminution de la quantité de fer intracellulaire par augmentation de son export. Cela confirme la fonction d’extrusion du fer intracellulaire de la ferro-GFP dans ce modèle. L’ajout d’hepcidine purifiée dans le milieu de culture induit une internalisation de la ferro-GFP, associée à une augmentation du fer intracellulaire. Cette action d’internalisation est rapide (2 à 3 heures) et s’observe pour des concentrations physiologiques d’hepcidine. De plus, cet effet est réversible car la ferro-GFP retrouve une localisation membranaire lorsque l’hepcidine est retirée du milieu. La même observation d’internalisation de la ferro-GFP est reproduite avec une hepcidine de 25 acides aminés synthétisée in vitro mais, de façon intéressante, pas avec la forme la plus courte de 20 acides aminés. Les auteurs montrent par des expériences d’immuno-marquage que la détection de ferro-GFP est diminuée en présence d’hepcidine et suggèrent sa dégradation via la voie lysosomale. La co-localisation de la ferro-GFP internalisée avec le marqueur lysosomal tardif Lamp1 renforce cette hypothèse. L’action est spécifique du couple hepcidine-ferroportine car l’internalisation de la ferro-GFP n’est pas observée en présence de protégrine, un autre peptide antimicrobien, et, inversement, l’hepcidine ne permet par l’internalisation du récepteur de l’EGF (epidermal growth factor). Les auteurs démontrent enfin, par des expériences d’immunoprécipitation, que la liaison entre l’hepcidine et la ferro-GFP est directe. Ainsi, trois ans après la démonstration du rôle crucial de l’hepcidine, ce travail permet-il de proposer une boucle de régulation assurant le contrôle de l’homéostasie du fer (Figure 1). Un excès de fer (par exemple, avec un régime riche en fer) induit l’expression hépatique d’hepcidine laquelle se lie directement avec la ferroportine pour entraîner son internalisation et sa dégradation. Le fer s’accumule alors dans la cellule et n’est plus exporté. La conséquence en est une diminution du transport du fer alimentaire au niveau des entérocytes matures. L’érythropoïèse qui doit être maintenue pour produire suffisamment de globules rouges pompe alors sur les réserves de fer en excès, ce qui conduit peu à peu à dépléter celles-ci et à rétablir un état d’équilibre. Du fait de sa petite taille, l’hepcidine a probablement une clairance très rapide, permettant ainsi une inversion rapide du processus.
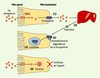 |
Figure 1. Modèle du mécanisme d’action de l’hepcidine. Après avoir été réduit, le fer alimentaire traverse la membrane apicale de l’entérocyte grâce au transporteur DMT1 puis transite vers la membrane basolatérale qu’il traverse en empruntant la ferroportine tout en étant réoxydé (1) (pour une revue plus détaillée des protéines impliquées, voir [1]). Le fer absorbé en excès est stocké au niveau du foie, provoquant une augmentation de la synthèse et de l’excrétion de l’hepcidine (par un mécanisme qui demeure inconnu) (2). L’hepcidine se lie alors directement sur la ferroportine (3), et induit son internalisation puis sa dégradation dans le lysosome (4). Le fer qui entre dans la cellule s’accumule alors dans la ferritine (5) et n’est plus exporté. Au final, le transport net du fer alimentaire est diminué et le pool de fer sera perdu lors de l’exfoliation de l’entérocyte au sommet de la villosité duodénale. Les réserves en fer (notamment hépatiques) sont alors utilisées pour maintenir une érythropoïèse efficace, ce qui, à terme, permet un retour à un état d’équilibre avec une diminution des réserves en fer. Lors d’une inflammation, une production excessive d’hepcidine pourrait aboutir à une dégradation trop importante de la ferroportine, engendrant une absorption insuffisante de fer. Un mécanisme similaire existerait au niveau des macrophages et des cellules du syncytiotrophoblaste, deux cibles tissulaires de l’hepcidine. |
Cet élégant travail, réalisé exclusivement sur un modèle cellulaire, demande maintenant une validation in vivo et pose la question de savoir si toutes les cellules exprimant la ferroportine vont être également sensibles à l’hepcidine. Outre l’entérocyte mature, l’hepcidine pourrait en effet agir sur l’export du fer, non seulement au niveau du système réticuloendothélial [2], mais également au niveau du placenta [5], en se liant sur la ferroportine qui est fortement exprimée dans les macrophages et les cellules du syncytiotrophoblaste placentaire. La prochaine étape concerne l’élucidation des interactions moléculaires entre l’hepcidine et la ferroportine et les mécanismes biochimiques mis en jeu lors de l’internalisation de la ferroportine. Il serait intéressant de déterminer à quel niveau (interaction et/ou internalisation) le processus est altéré avec les formes mutées de l’hepcidine (mutations faux-sens) identifiées chez l’homme dans des surcharges en fer sévères. De la même façon, le rôle du peptide de 20 acides aminés reste indéterminé. S’agit-il uniquement d’un produit de dégradation de la forme de 25 acides aminés ?
D’après les auteurs, la ferroportine représente le « récepteur » de l’hepcidine, mais l’hepcidine a-t-elle livré tous ses secrets ? Il est en effet légitime de se demander si l’hepcidine ne pourrait pas agir (suivant sa concentration, ses partenaires, sa cible…) en utilisant un autre procédé.
Quoi qu’il en soit, ce travail constitue une étape décisive pour le développement de molécules mimant les effets de l’hepcidine sur l’internalisation de la ferroportine pour traiter des situations physiopathologiques où l’hormone est trop faiblement produite (hémochromatoses et autres). À l’inverse, le développement de molécules qui s’opposent à l’action de l’hepcidine sera recherché pour traiter des situations où la concentration urinaire de l’hormone est trop élevée, comme c’est le cas lors d’épisodes inflammatoires ou infectieux [7, 8]. L’hepcidine en excès entraînant alors une dégradation trop importante de la ferroportine pourrait expliquer la mise en place de l’anémie fréquemment observée dans les états inflammatoires chroniques.
Références
- Beaumont C. Mécanismes moléculaires de l’homéostasie du fer. Med Sci (Paris) 2004; 20 : 68–72. [Google Scholar]
- Nicolas G, Benounn M, Devaux I, et al. Lack of hepcidin gene expression and severe tissue iron overload in upstream stimulatory factor (USF2) knockout mice. Proc Natl Acad Sci USA 2001; 98 : 8780–5. [Google Scholar]
- Pigeon C, Ilyin G, Courselaud B, et al. A new mouse liver-specific gene, encoding a protein homologous to human antimicrobial peptide hepcidin, is overexpressed during iron overload. J Biol Chem 2001; 276 : 7811–9. [Google Scholar]
- Roetto A, Papanikolaou G, Politou M, et al. Mutant antimicrobial peptide hepcidin is associated with severe juvenile hemochromatosis. Nat Genet 2002; 33 : 21–2. [Google Scholar]
- Nicolas G, Benounn M, Porteu A, et al. Severe iron deficiency anemia in transgenic mice expressing liver hepcidin. Proc Natl Acad Sci USA 2002; 99 : 4596–601. [Google Scholar]
- Nemeth E, Tuttle MS, Powelson J, et al. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science Express 28 octobre 2004. [Google Scholar]
- Nicolas G, Chauvet C, Viatte L, et al. The gene encoding the iron regulatory peptide hepcidin is regulated by anemia, hypoxia, and inflammation. J Clin Invest 2002; 110 : 1037–44. [Google Scholar]
- Nemeth E, Valore EV, Territo M, et al. Hepcidin, a putative mediator of anemia of inflammation, is a type II acute-phase protein. Blood 2003; 101 : 2461–3. [Google Scholar]
© 2005 médecine/sciences - Inserm / SRMS
Liste des figures
|
Figure 1. Modèle du mécanisme d’action de l’hepcidine. Après avoir été réduit, le fer alimentaire traverse la membrane apicale de l’entérocyte grâce au transporteur DMT1 puis transite vers la membrane basolatérale qu’il traverse en empruntant la ferroportine tout en étant réoxydé (1) (pour une revue plus détaillée des protéines impliquées, voir [1]). Le fer absorbé en excès est stocké au niveau du foie, provoquant une augmentation de la synthèse et de l’excrétion de l’hepcidine (par un mécanisme qui demeure inconnu) (2). L’hepcidine se lie alors directement sur la ferroportine (3), et induit son internalisation puis sa dégradation dans le lysosome (4). Le fer qui entre dans la cellule s’accumule alors dans la ferritine (5) et n’est plus exporté. Au final, le transport net du fer alimentaire est diminué et le pool de fer sera perdu lors de l’exfoliation de l’entérocyte au sommet de la villosité duodénale. Les réserves en fer (notamment hépatiques) sont alors utilisées pour maintenir une érythropoïèse efficace, ce qui, à terme, permet un retour à un état d’équilibre avec une diminution des réserves en fer. Lors d’une inflammation, une production excessive d’hepcidine pourrait aboutir à une dégradation trop importante de la ferroportine, engendrant une absorption insuffisante de fer. Un mécanisme similaire existerait au niveau des macrophages et des cellules du syncytiotrophoblaste, deux cibles tissulaires de l’hepcidine. |
| Dans le texte | |
Les statistiques affichées correspondent au cumul d'une part des vues des résumés de l'article et d'autre part des vues et téléchargements de l'article plein-texte (PDF, Full-HTML, ePub... selon les formats disponibles) sur la platefome Vision4Press.
Les statistiques sont disponibles avec un délai de 48 à 96 heures et sont mises à jour quotidiennement en semaine.
Le chargement des statistiques peut être long.