")
")
| Issue |
Med Sci (Paris)
Volume 18, Number 8-9, Août–Septembre 2002
|
|
|---|---|---|
| Page(s) | 802 - 803 | |
| Section | Le Magazine : Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/20021889802 | |
| Published online | 15 août 2002 | |
La somatostatine, peptide moteur de la migration neuronale ?
Involvement of somatostatin in migratory locomotion of neurons
1
Inserm U.549, IFR Broca-Sainte Anne, 2 ter, rue d’Alésia, 75014 Paris, Fran
2
Inserm U.413, IFRMP 23, Laboratoire de neuroendocrinologie cellulaire et moléculaire, Université de Rouen, 76821 Mont-Saint-Aignan Cedex, Fran
Les neuropeptides exercent leurs fonctions multiples dans le cerveau en agissant par l’intermédiaire de récepteurs spécifiques. Dans la plupart des régions centrales, les systèmes peptidergiques se mettent en place progressivement au cours de l’ontogenèse selon un schéma comparable à celui que l’on trouve chez l’adulte. Dans certaines structures centrales, cependant, des peptides et leurs récepteurs sont détectés de manière précoce et transitoire, suggérant leur rôle à certaines étapes bien précises du développement [1].
La somatostatine est un neuropeptide largement exprimé dans le système nerveux central et qui intervient dans de multiples fonctions cérébrales par l’intermédiaire de cinq récepteurs (sst1-sst5) couplés aux protéines Gi/Go [2]. Elle est transitoirement mise en évidence à des stades-clés de la maturation dans plusieurs régions centrales comme le cervelet, la moelle épinière, le cortex ou le système visuel [1]. Des récepteurs fonctionnels du peptide sont détectés simultanément dans ces mêmes régions [3], en particulier au niveau de neurones immatures.
Le cervelet de rongeur est apparu très vite comme un modèle de choix pour étudier in vitro les mécanismes cellulaires induits par le peptide in vivo. La somatostatine est en effet fortement et transitoirement détectée dans les cellules de Golgi et de Purkinje du cervelet lors de son développement au cours des trois premières semaines de vie post-natale. Son expression diminue par la suite et, chez l’adulte, est restreinte à de rares cellules [1]. À la même période, une forte liaison somatostatinergique est détectée au niveau du neuro-épithélium germinatif secondaire à l’origine des cellules en grains, la couche granulaire externe (CGE) (Figure 1) [4]. Ces sites de liaison spécifiques sont associés aux précurseurs des neurones en grains, ils sont fonctionnels in vitro [5], et leur caractérisation pharmacologique et moléculaire a permis de conclure qu’ils correspondent essentiellement à des récepteurs sst2 même si une faible proportion de sites de type sst1 est toutefois détectable [6].
 |
Figure 1. Illustration de l’expression transitoire des sites de liaison de la somatostatine visualisés par autoradiographie au cours du développement du cortex cérébelleux des rongeurs. Les flèches rouges indiquent le marquage associé à la couche granulaire externe. 4J-12J : 4 à 12 jours de vie post-natale ; Ad : stade adulte. Barre d’échelle : 5 mm (modifiée d’après [4]). |
La fonction de la somatostatine dans le cervelet immature était, jusqu’à il y a peu, mal définie : in vitro, le peptide stimulait modérément la formation de neurites et l’expression de marqueurs de différenciation sur des cellules granulaires, mais ne semblait influencer ni la prolifération ni la survie des neuroblastes. Récemment, l’enregistrement en temps réel de cellules granulaires marquées à l’aide d’une sonde fluorescente a montré que la somatostatine est un puissant régulateur de la migration neuronale dans des explants cérébelleux [7]. L’ajout de somatostatine dans le milieu de culture accélère la migration tangentielle des cellules granulaires dans la CGE, couche d’où elles sont issues, tandis qu’elle la diminue légèrement dans la couche moléculaire, et fortement dans la CGI qui constitue leur destination finale (Figure 2). Le peptide endogène est également impliqué dans ces effets puisque l’incubation de tranches de tissu de cette région en présence d’un antagoniste de la somatostatine entraîne les effets inverses. Le peptide semble agir directement sur les cellules granulaires, puisqu’il exerce des effets similaires sur des neuroblastes granulaires isolés provenant d’explants cérébelleux : il active le mouvement de cellules immatures (1 jour de culture) et ralentit celui de cellules plus différenciées (2 jours de culture). Les différences d’effet du peptide sur la vitesse de migration cellulaire semblent donc liées au stade de maturation de la cellule. La somatostatine affecte sélectivement la vitesse de migration de la cellule puisque ni la longueur des neurites, ni la forme de la cellule, ni la direction et le mode saltatoire de déplacement ne sont modifiés dans ce modèle. En parallèle, la somatostatine régularise et augmente les oscillations calciques dans les cellules immatures ; elle élimine les courants transitoires calciques établis à un stade plus différencié, par l’intermédiaire de canaux calciques ou potassiques, effecteurs du peptide, comme cela a été démontré dans d’autres modèles [2]. Les actions positive et négative du peptide sur la migration des cellules granulaires pourraient résulter de l’activation successive ou simultanée de différents sous-types de récepteurs au cours de la maturation des cellules en grains puisqu’un tel antagonisme des récepteurs sst2 et sst1 sur les courants AMPA1 glutamatergiques a été précédemment démontré dans des cultures neuronales [6].
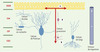 |
Figure 2. Représentation schématique des effets de la somatostatine sur la migration des cellules granulaires du cortex cérébelleux de souris âgée de 10 jours. Les flèches rouges indiquent les sens de migration tangentielle (1), radiale dépendante de la glie (2) et radiale indépendante de la glie (3). Les neurones exprimant la somatosta-tine sont représentés par un corps cellulaire coloré en bleu. La cellule en grains, à différents stades de développement, est représentée en rose. La colonne de droite visualise l’action stimulatrice (+), modérément inhibitrice (-) et fortement inhibitrice (—) de la somatostatine sur la migration neuronale. CGE : couche granulaire externe ; CGI : couche granulaire interne ; CM : couche moléculaire ; CP : couche des cellules de Purkinje. |
L’étude récente d’animaux génétiquement modifiés pour la somatostatine ou pour le récepteur sst2 conforte l’implication du peptide dans la mise en place des réseaux cérébelleux. Les performances des animaux témoins et mutants sont voisines dans plusieurs tests de motricité et de coordination, démontrant l’absence de défaut moteur majeur chez l’adulte [8, 9]. Néanmoins, un déficit notable de l’apprentissage moteur est décelé chez des souris dont le gène codant pour la somatostatine a été invalidé lorsque la demande au niveau de la coordination motrice est augmentée, mettant ainsi en évidence un phénotype subtil qui pourrait être associé à un défaut de maturation du cervelet [9].
La somatostatine apparaît donc désormais comme un neuropeptide important dans le contrôle de la migration des cellules granulaires du cervelet. Cette activité de la somatostatine pourrait refléter une fonction plus générale concernant l’ensemble du système nerveux central en développement, mais aussi d’autres modèles où la migration cellulaire intervient, comme l’hématopoïèse [10].
α-amino-3-hydroxy-5-méthyl-4isoxazole-proprionate.
Références
- Shiosaka S. Ontogeny of the central somatostatinergic system. In : Björklund A, Hökfelt T, Tohyama M, eds. Handbook of chemical neuroanatomy. Amsterdam : Elsevier, 1992: 369–98. [Google Scholar]
- Reisine T, Bell GI. Molecular biology of somatostatin receptors. Endocrinol Rev 1995; 16 : 427–42. [Google Scholar]
- Gonzalez BJ, Leroux P, Bodenant C, Laquerriere A, Coy DH, Vaudry H. Ontogeny of somatostatin receptors in the rat brain. Neuroscience 1989; 29 : 629–44. [Google Scholar]
- Gonzalez BJ, Leroux P, Bodenant C, Braquet P, Vaudry H. Pharmacological characterization of somatostatin receptors in the rat cerebellum during development. J Neurochem 1990; 55 : 154–7. [Google Scholar]
- Gonzalez B, Leroux P, Lamacz M, Bodenant C, Balazs R, Vaudry H. Somatostatin receptors are expressed by immature cerebellar granule cells: evidence for a direct inhibitory effect of somatostatin on neuroblast activity. Proc Natl Acad Sci USA 1992; 89 : 9627–31. [Google Scholar]
- Lanneau C, Viollet C, Faivre-Bauman A, et al. Somatostatin receptor subtypes sst1 and sst2 elicit opposite effects on the response to glutamate of mouse hypothalamic neurones: an electrophysiological and single cell RT-PCR study. Eur J Neurosci 1998; 10 : 204–12. [Google Scholar]
- Yacubova E, Komuro H. Stage-specific control of neuronal migration by somatostatin. Nature 2002; 3 : 77–81. [Google Scholar]
- Viollet C, Vaillend C, Videau C, et al. Involvement of sst2 somatostatin receptor in locomotor, exploratory activity and emotional reactivity in mice. Eur J Neurosci 2000; 12 : 3761–70. [Google Scholar]
- Zeyda T, Diehl N, Paylor R, Brennan MB, Hochgeschwender U. Impairment in motor learning of somatostatin null mutant mice. Brain Res 2001; 906 : 107–14. [Google Scholar]
- Oomen SP, van Hennik PB, Antonissen C, et al. Somatostatin is a selective chemoattractant for primitive (CD34+) hematopoietic progenitor cells. Exp Hematol 2002; 30 : 116–25. [Google Scholar]
© 2002 médecine/sciences - Inserm / SRMS
Liste des figures
|
Figure 1. Illustration de l’expression transitoire des sites de liaison de la somatostatine visualisés par autoradiographie au cours du développement du cortex cérébelleux des rongeurs. Les flèches rouges indiquent le marquage associé à la couche granulaire externe. 4J-12J : 4 à 12 jours de vie post-natale ; Ad : stade adulte. Barre d’échelle : 5 mm (modifiée d’après [4]). |
| Dans le texte | |
|
Figure 2. Représentation schématique des effets de la somatostatine sur la migration des cellules granulaires du cortex cérébelleux de souris âgée de 10 jours. Les flèches rouges indiquent les sens de migration tangentielle (1), radiale dépendante de la glie (2) et radiale indépendante de la glie (3). Les neurones exprimant la somatosta-tine sont représentés par un corps cellulaire coloré en bleu. La cellule en grains, à différents stades de développement, est représentée en rose. La colonne de droite visualise l’action stimulatrice (+), modérément inhibitrice (-) et fortement inhibitrice (—) de la somatostatine sur la migration neuronale. CGE : couche granulaire externe ; CGI : couche granulaire interne ; CM : couche moléculaire ; CP : couche des cellules de Purkinje. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.