")
")
| Issue |
Med Sci (Paris)
Volume 42, Number 6-7, Juin-Juillet 2026
|
|
|---|---|---|
| Page(s) | 544 - 547 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/2026097 | |
| Published online | 17 juillet 2026 | |
Comment redessiner la carte du cerveau en écoutant les neurones un par un ?
How to redraw the brain’s map by listening to the activity of single neurons?
1
Laboratoire plasticité du cerveau, École supérieure de physique et de chimie industrielles de la ville de Paris (ESPCI Paris), Université Paris sciences et lettres, CNRS UMR 8249, Paris, France
2
Centre de recherche en neurosciences de Lyon, CNRS UMR 5292, Inserm U1028, Lyon, France
3
Department of neuroscience, Karolinska Institutet, Stockholm, Suède
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
**
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Pour s’orienter dans le cerveau, les neuroscientifiques utilisent des cartes anatomiques. Ces cartes sont contenues dans des atlas de référence et ont été établies historiquement en observant la forme et la disposition des cellules neuronales par microscopie. Cette tradition, vieille de plus d’un siècle, permet de diviser le cerveau en régions distinctes. Mais à quel point ces divisions sont- elles pertinentes pour comprendre son fonctionnement ? Une étude réalisée chez la souris remet en question les divisions anatomiques traditionnelles en enregistrant l’activité cérébrale neurone par neurone [1].
D’où viennent les cartes du cerveau ?
L’histoire de la cartographie du cerveau est intimement liée à l’histoire des neurosciences. Dès l’origine, il est très difficile de séparer l’étude des fonctions cérébrales de l’anatomie du cerveau car les neurosciences prennent leurs racines dans des disciplines étudiant fonction et anatomie côte à côte, comme la physiologie ou l’histologie, mais aussi dans la neuropsychiatrie et la psychologie [m/s 2]. Dès 1776, Francesco Gennari découvre dans le néocortex une strie visible à l’œil nu qui correspond à la couche 4 du cortex visuel primaire des primates (ainsi appelé cortex strié), et qui permet de séparer cette aire des aires corticales voisines. Au début du XIXe siècle, l’anatomiste et physiologiste allemand Franz Joseph Gall (1758-1828) propose que les capacités et qualités de chaque individu puissent être identifiées par un examen de la forme du crâne (Figure 1A). De cette discipline pseudoscientifique nommée phrénologie1 [3], découle le « localisationnisme », dont l’objectif est d’associer des fonctions sensorielles et motrices à des aires cérébrales spécialisées (voir notamment les travaux du médecin et anatomiste français Paul Broca (1824-1880), et la découverte des aires du langage [4]). Le projet de cartographie du cerveau humain est lancé. À la fin du XIXe siècle, la théorie neuronale marque un tournant majeur dans les neurosciences. Formulée notamment par Santiago Ramón y Cajal (1852-1934) et Wilhelm Waldeyer (1836-1921), cette théorie postule que le neurone est l’unité de base du système nerveux, à la fois sur le plan structurel (une cellule différenciée, dotée d’un corps cellulaire, d’un axone et de dendrites) et sur le plan fonctionnel (une entité capable de recevoir, d’intégrer et de transmettre des signaux de nature électrique) [5]. Cette avancée conceptuelle, en affirmant que le cerveau est composé de cellules individuelles et interconnectées plutôt que d’un réseau continu (comme le suggéraient les tenants de la théorie réticulaire), a transformé la manière d’étudier le système nerveux. C’est dans ce contexte, à Berlin, alors épicentre des recherches en neuroanatomie, que cette théorie a inspiré une nouvelle génération de cartographes du cerveau. En 1900, Cécile (1875-1962) et Oskar (1870-1959) Vogt fondent à Berlin un institut de recherche privé (Neurologisches Laboratorium), avec un objectif précis : établir une carte du cortex cérébral en utilisant les techniques les plus avancées de l’époque : la cytoarchitectonie ou observation du nombre et de la forme des cellules par microscopie, la myéloarchitectonie ou analyse de l’arrangement des fibres nerveuses, l’étude de l’impact fonctionnel des lésions du cortex cérébral, et les stimulations électriques du cerveau durant les interventions neurochirurgicales (travaux de l’américain Wilder Penfield (1891-1976) [6]). Pourtant, c’est un autre chercheur de cet institut, Korbinian Brodmann (1868-1918), qui s’illustre au sein de la communauté scientifique en proposant, en 1909, une division du cortex cérébral en 50 régions (les époux Vogt, quant à eux, en trouvèrent 200) (Figure 1B), uniquement fondée sur la cytoarchitectonie, et la carte qu’il a établie est celle encore utilisée aujourd’hui [7]. La carte établie par Cécile et Oskar Vogt, pourtant plus complète car intégrant des données de connectivité et des résultats d’expériences fonctionnelles [8, 9], n’accèdera pas à la même notoriété que celle de Brodmann.
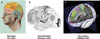 |
Figure 1 Représentations successives du cerveau humain illustrant l’évolution des hypothèses et des méthodes de cartographie cérébrale : de la phrénologie aux atlas modernes. A. Hypothèses localisatrices sous-jacentes à la phrénologie, où les fonctions cérébrales étaient associées à des bosses crâniennes (© Nirupa Rao). B. Découpage cytoarchitectonique du cortex cérébral par Brodmann, fondé sur l’organisation cellulaire et toujours utilisé comme référence anatomique (adapté de [3]). C. Projets collaboratifs contemporains à grande échelle, comme eBRAINS, offrant une modélisation tridimensionnelle du cerveau humain fondée sur la cytoarchitecture, avec des aires cérébrales interactives. |
Au cours de la dernière décennie, la communauté des neurosciences s’est lancée dans des projets d’envergure colos-sale pour cartographier les cerveaux de plusieurs espèces animales. L’approche utilisée est similaire à celle des époux Vogt : utiliser les techniques de pointe cellulaires et moléculaires pour faire l’inventaire des composants du cerveau et tenter d’en expliquer le fonctionnement. Aux États-Unis, c’est le projet BRAIN2, soutenu financièrement par l’institut Allen et la fondation Kavli, qui produit des atlas du cerveau avec un niveau de précision sans précédent [10]. En Europe, le Human brain project a abouti à la création de la plateforme eBRAINS3. Cette infrastructure, ouverte à la communauté scientifique internationale, propose des atlas du cerveau de haute résolution (comme l’atlas Julich-Brain ou l’atlas BigBrain) (Figure 1C), ainsi que des outils pour visualiser, analyser et modéliser les données cérébrales à différentes échelles, du neurone individuel aux réseaux neuronaux. Les « aires de Brodmann », définissant les « organes du cerveau », servent toujours de toile de fond à ces atlas.
Comprendre la fonction des régions du cerveau
En biologie, la fonction est intimement liée à la structure. Selon ce principe, les régions du cerveau ayant des agencements cellulaires différents doivent correspondre à des fonctions différentes. Les connexions de certaines régions avec les organes sensoriels ou avec les muscles squelettiques ont permis de préciser ces fonctions. Ainsi, la région du cortex connectée à la rétine correspond à l’aire « visuelle », et une région qui active un ensemble de muscles est une aire « motrice ». Mais cela se complique lorsque l’on s’intéresse à des processus mentaux plus abstraits, comme la pensée, même si Brodmann était convaincu que son découpage du cortex cérébral en aires permettrait d’identifier des « organes de la pensée ». Plus concrètement, comment définir une région cérébrale qui permet de prendre des décisions ou de planifier une action ?
La méthode qui a prouvé son efficacité pour comprendre la fonction d’une région du cerveau repose sur deux principes : 1) La région A est-elle active quand je réalise la fonction X ? 2) En désactivant la région A, perturbe-t-on la fonction X ? Si un individu présente une lésion dans la région A du cortex cérébral, et que cette lésion l’empêche de prononcer autre chose que le son tan, alors cette aire est certainement impliquée dans le langage (cf. les travaux de Paul Broca [4]). C’est ainsi que, depuis les premiers travaux des phrénologistes au début du XIXe siècle jusqu’à l’avènement de techniques avancées d’imagerie cérébrale comme l’imagerie par résonance magnétique à ultra-haute résolution [11], les neuroscientifiques progressent dans la cartographie des fonctions régionales du cerveau. Néanmoins, ils se sont vite rendu compte que les aires cérébrales sont « multitâches » (bien que ce constat fasse encore l’objet d’un débat scientifique [12]). Lorsque l’activité neuronale du cortex préfrontal, une région située à l’avant du cerveau, est perturbée chez un individu, on constate un déficit de planification et d’organisation des actions, des difficultés à prendre des décisions, des troubles de l’attention et de la concentration, ainsi que des réactions émotionnelles excessives et une perte de motivation [13, 14]. Face à cette multiplicité des « fonctions » associées à une même région cérébrale, les neuroscientifiques ont compris qu’il était plus pertinent d’étudier non pas des aires cérébrales isolées, mais des réseaux associant plusieurs aires qui interagissent et collaborent pour accomplir une fonction mentale donnée.
Récemment, un consortium de 14 laboratoires, l’International brain laboratory, a davantage remis en cause cet axiome reliant la structure à la fonction [15]. Dans un projet rendu possible par les progrès techniques, ce consortium a cherché à déterminer les régions du cerveau possédant des neurones actifs quand les animaux sont en mouvement. Pour cela, sans idée préconçue quant à la localisation de l’activité neuronale liée à la motricité, les chercheurs ont enregistré simultanément l’activité de neurones uniques dans un grand nombre de structures cérébrales. À leur grande surprise, toutes les régions du cerveau dont l’activité a pu être enregistrée possèdent des neurones actifs pendant le mouvement. Ce résultat n’est pas sans rappeler les hypothèses anti-localisationnistes de Karl Lashley (1890-1958) [16]. Remetil pour autant en cause la pertinence de la régionalisation du cerveau, et notamment la cartographie corticale de Brodmann ? Il importe de souligner que ce débat sur la relation entre structure et fonction repose souvent sur une interprétation réductrice des travaux de Brodmann, dont l’approche scientifique, parfois caricaturée comme définissant des régions corticales « rigides », était en réalité plus nuancée. Dans une partie moins citée de son ouvrage, Brodmann lui-même précise que « [...] il est difficile de concevoir l’émergence de [processus cognitifs] d’une autre manière qu’au travers d’infinies et complexes interactions de nombreux processus élémentaires, c’est-à dire du fonctionnement simultané d’autant d’aires corticales que le cortex en compte, voire de régions sous-corticales » [7]. Ainsi, loin de défendre une segmentation figée du cerveau, Brodmann avait déjà l’idée de son organisation dynamique et distribuée.
Une cartographie qui utilise le langage des neurones
Le cortex préfrontal, une des régions les plus énigmatiques du cerveau, serait responsable de la grande majorité des fonctions cognitives de haut niveau comme l’attention, la prise de décision, la planification, et la perception consciente. On considère aussi que son rôle est de structurer le présent pour planifier le futur [14, 17]. L’hypothèse souvent avancée est que les différentes sous-régions qui le composent reflètent des spécialisations fonctionnelles distinctes. Cependant, cette vision parcellaire du cortex préfrontal, bien qu’historiquement fondatrice, se heurte aujourd’hui aux limites d’une approche purement anatomique : d’où l’émergence de stratégies innovantes, comme celle développée par l’équipe dirigée par Marie Carlén à Stockholm (Suède), qui propose de repenser cette cartographie à partir de l’activité même des neurones.
Afin de créer une nouvelle carte du cortex préfrontal de la souris fondée sur l’activité des neurones individuels (Figure 2A), ces chercheurs ont utilisé des sondes Neuropixels, une technique permettant d’enregistrer simultanément l’activité de centaines de neurones individuels dans plusieurs régions cérébrales. Grâce à ces sondes, positionnées avec précision par stéréotaxie à l’aide d’un atlas anatomique (Figure 2B), ils ont cartographié en trois dimensions l’emplacement et l’activité de plus de 24 000 neurones dans le cortex préfrontal [1]. Les neurones communiquent entre eux en émettant des signaux électriques appelés « potentiels d’action ». Les chercheurs ont analysé la fréquence et le rythme d’émission des potentiels d’actions lorsque les souris ne réalisent aucune tâche particulière. En identifiant des groupes de neurones présentant des profils d’activité similaires, ils ont pu observer dans quels territoires ces neurones étaient surreprésentés ou absents. Cette analyse a révélé une nouvelle carte du cortex préfrontal, radicalement différente de celle, fondée sur l’anatomie, des atlas existants (Figure 2C, 2D). Peu de temps auparavant, le groupe dirigé par Jun Yan, à Shanghai (Chine), avait publié un article concernant la connectivité des neurones, également dans le cortex préfrontal de la souris [18]. Ensemble, ces deux études montrent que les territoires fonctionnels reflètent l’organisation des réseaux neuronaux, c’est-à-dire la manière dont les neurones sont connectés entre eux. L’organisation de cette connectivité est complexe, mais on peut l’imager comme des axes routiers qui contraignent la circulation de l’information neuronale. Cette circulation suit une organisation hiérarchique : certains territoires, souvent situés « en amont » dans la hiérarchie, reçoivent et intègrent des informations de multiples sources avant d’en envoyer vers d’autres régions, tandis que les territoires « en aval » sont davantage spécialisés dans le traitement et la transmission d’informations spécifiques vers les niveaux hiérarchiques supérieurs. Ici, le lien est établi entre cette hiérarchie de communication et le « langage » des neurones. Par exemple, les neurones qui émettent des potentiels d’action réguliers avec une fréquence faible se situent dans des territoires en amont dans la hiérarchie. L’activité d’un tel neurone permet une intégration de l’information sur le long terme, et cette propriété est associée à des fonctions cognitives comme la mémoire de travail et la planification de l’action.
 |
Figure 2 Cartographie fonctionnelle du cortex préfrontal de la souris fondée sur l’activité de neurones individuels. A. Activité de neurones individuels enregistrée à l’aide de sondes Neuropixels. La fréquence et la régularité ou non des potentiels d'action permet de distinguer deux principaux types de neurones, représentés ici en bleu et en rouge. B. Positionnement des neurones enregistrés dans un atlas tridimensionnel du cerveau murin. C. Passage en deux dimensions (carte aplatie schématique) D. Cartographie fonctionnelle, révélant de nouveaux territoires définis par les profils d’activité neuronale. |
C’est d’ailleurs ce que les recherches de l’équipe dirigée par Marie Carlén ont pu mettre en évidence. En réanalysant les données de l’étude du International brain laboratory [15], cette équipe a découvert que les territoires du cortex préfrontal identifiés à partir de l’activité neuronale lorsque les souris sont « au repos » coïncident avec ceux majoritairement activés lors des processus de prise de décision chez ces animaux. Cette correspondance suggère que les profils d’activité des neurones lorsque la souris est éveillée mais au repos – considérés à tort comme un « bruit de fond » – pourraient en réalité refléter une organisation fonctionnelle sous- jacente, préfigurant les réseaux neuronaux recrutés pendant les tâches cognitives.
Un principe pour le futur ?
Cette approche de cartographie fonctionnelle, fondée sur l’activité et les connections des neurones, ouvre la voie à de nouvelles études utilisant le même principe de cartographie dans d’autres parties du cerveau ou chez d’autres espèces que la souris. En dépassant les frontières anatomiques traditionnelles, une telle approche permet d’envisager les régions cérébrales, non plus comme des entités statiques fixant la fonction des neurones, mais comme des entités abritant des réseaux dynamiques dont les propriétés émergent de la connectivité. Il devient ainsi possible de déterminer le rôle de la distribution de l’activité neuronale dans de multiples aires corticales lors d’un mouvement ou pendant un processus cognitif.
Cette nouvelle approche de la cartographie à l’échelle cellulaire se développe au moment où des cartographies moléculaires (comme celle fondée sur l’analyse des profils d’expression génique à l’échelle de la cellule unique) viennent compléter ces approches anatomiques et fonctionnelles. Ainsi, les résultats d’études de « transcriptomique spatiale » (comme celles menées dans le cadre du Human cell atlas ou du Mouse brain atlas) ont révélé que des populations neuronales ayant des profils d’expression génique similaires peuvent s’organiser en réseaux fonctionnels cohérents même si elles sont réparties dans des régions anatomiques distinctes [19]. Ainsi, il est probable que l’intégration de ces données multi-échelles – structure, connectivité, activité, et profil moléculaire – offrira une compréhension sans précédent de la manière dont le cerveau exécute ses fonctions.
Ces nouvelles approches cellulaires et moléculaires remettent en question le modèle d’une division du travail entre les différentes régions au profit d’un modèle plus intégré du fonctionnement cérébral, mais la cartographie du cortex cérébral, dont Brodmann et les époux Vogt ont été les pionniers, restera encore longtemps utile comme cadre des discussions neuroscientifiques.
Liens d’intérêt
Les auteurs déclarent ne pas avoir de lien d’intérêt.
Références
- Le Merre P, Heining K, Slashcheva M, et al. A prefrontal cortex map based on single-neuron activity. Nat Neurosci 2026 ; 29 : 673–81. [Google Scholar]
- Paillette C, Griset P, Agid Y. Aux origines des neurosciences - L’héritage et les fondations, du XIXe siècle aux années 1940. Med Sci (Paris) 2021 ; 37 : 793–8. [Google Scholar]
- Poskett J. Materials of the mind: Phrenology, race, and the global history of science, 1815-1920. Chicago, IL : University of Chicago Press, 2022 : 373 p. [Google Scholar]
- Broca P. Remarques sur le siège de la faculté du langage articulé, suivies d’une observation d’aphémie (perte de la parole). Bull Soc Anat Paris 1861 ; 36 : 330–57. [Google Scholar]
- Guillery RW. Observations of synaptic structures: origins of the neuron doctrine and its current status. Philos Trans R Soc Lond B Biol Sci 2005 ; 360 : 1281–307. [Google Scholar]
- Penfield W, Boldrey E. Somatic motor and sensory representation in the cerebral cortex of man as studied by electrical stimulation. Brain 1937 ; 60 : 389–443. [Google Scholar]
- Brodmann K. Vergleichende Lokalisationslehre der Grosshirnrinde in ihren Prinzipien dargestellt auf Grund des Zellenbaues. 1909. [Google Scholar]
- Vogt C, Vogt O. Die vergleichend-architektonische und die vergleichend-reizphysiologische Felderung der Großhirnrinde unter besonderer Berücksichtigung der menschlichen. Naturwissenschaften 1926 ; 14 : 1190–4. [Google Scholar]
- Pessoa L. The entangled brain: How perception, cognition, and emotion are woven together. The MIT Press, 2022. [Google Scholar]
- Yao Z, van Velthoven CTJ, Kunst M, et al. A high-resolution transcriptomic and spatial atlas of cell types in the whole mouse brain. Nature 2023 ; 624 : 317–32. [Google Scholar]
- Ramos-Llordén G, Lee H-H, Davids M, et al. Ultra-high gradient connectomics and microstructure MRI scanner for imaging of human brain circuits across scales. Nat Biomed Eng 2026 ; 10 : 309–24. [Google Scholar]
- Fodor JA. Modularité de l’esprit. Paris : MINUIT, 1986 : 176 p. [Google Scholar]
- Damasio H, Grabowski T, Frank R, et al. The return of Phineas Gage: Clues about the brain from the skull of a famous patient. Science 1994 ; 264 : 1102–5. [Google Scholar]
- Fuster JM. The prefrontal cortex, 5th edition. Academic Press. 2015. [Google Scholar]
- International Brain Laboratory, Angelaki D, Benson B, et al. A brain-wide map of neural activity during complex behaviour. Nature 2025 ; 645 : 177–91. [Google Scholar]
- Lashley KS. In search of the engram. Physiological mechanisms in animal behavior. (Society’s Symposium IV.). Oxford, England : Academic Press, 1950 : 454–82. [Google Scholar]
- Le Merre P, Ährlund-Richter S, Carlén M. The mouse prefrontal cortex: Unity in diversity. Neuron 2021 ; 109 : 1925–44. [Google Scholar]
- Gao L, Liu S, Gou L, et al. Single-neuron projectome of mouse prefrontal cortex. Nat Neurosci 2022 ; 25 : 515–29. [Google Scholar]
- Hayden BY, Heilbronner SR, Yoo SBM. Rethinking the centrality of brain areas in understanding functional organization. Nat Neurosci 2026 ; 29 : 267–78. [Google Scholar]
Terme dérivé du grec ancien phren, qui désigne le cerveau.
Acronyme de « brain research through advancing innovative neurotechnologies » (https://braininitiative.nih.gov/).
© 2026 médecine/sciences – Inserm
 Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l’utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Article publié sous les conditions définies par la licence Creative Commons Attribution License CC-BY (https://creativecommons.org/licenses/by/4.0), qui autorise sans restrictions l’utilisation, la diffusion, et la reproduction sur quelque support que ce soit, sous réserve de citation correcte de la publication originale.
Liste des figures
|
Figure 1 Représentations successives du cerveau humain illustrant l’évolution des hypothèses et des méthodes de cartographie cérébrale : de la phrénologie aux atlas modernes. A. Hypothèses localisatrices sous-jacentes à la phrénologie, où les fonctions cérébrales étaient associées à des bosses crâniennes (© Nirupa Rao). B. Découpage cytoarchitectonique du cortex cérébral par Brodmann, fondé sur l’organisation cellulaire et toujours utilisé comme référence anatomique (adapté de [3]). C. Projets collaboratifs contemporains à grande échelle, comme eBRAINS, offrant une modélisation tridimensionnelle du cerveau humain fondée sur la cytoarchitecture, avec des aires cérébrales interactives. |
| Dans le texte | |
|
Figure 2 Cartographie fonctionnelle du cortex préfrontal de la souris fondée sur l’activité de neurones individuels. A. Activité de neurones individuels enregistrée à l’aide de sondes Neuropixels. La fréquence et la régularité ou non des potentiels d'action permet de distinguer deux principaux types de neurones, représentés ici en bleu et en rouge. B. Positionnement des neurones enregistrés dans un atlas tridimensionnel du cerveau murin. C. Passage en deux dimensions (carte aplatie schématique) D. Cartographie fonctionnelle, révélant de nouveaux territoires définis par les profils d’activité neuronale. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.