")
")
Figure 5.
Télécharger l'image originale
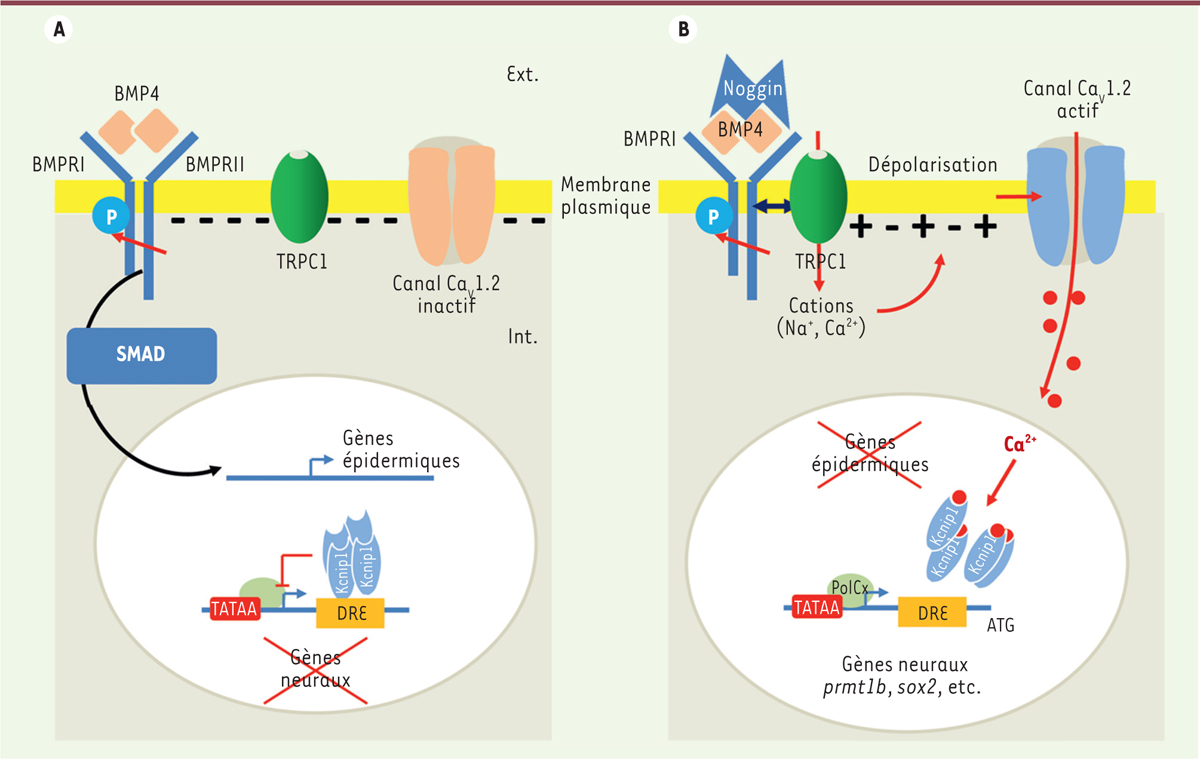
Les acteurs identifiés de la transduction du signal neuralisant. A. La signalisation BMP4 met en jeu des récepteurs à activité sérine/thréonine kinase (BMPRI et BMPRII). Lors de la fixation de BMP4 sous forme de dimères, la phosphorylation de BMPRI par BMPRII déclenche la phosphorylation des protéines effectrices SMAD et leur relocalisation dans le noyau où elles agissent alors comme facteurs de transcription sur les gènes cibles (gènes épidermiques). Lorsque la voie BMP est active, les gènes neuraux sont réprimés. Dans l’embryon, ce mécanisme est actif dans l’ectoderme ventral, destiné à donner du tissu épidermique. B. La protéine extracellulaire Noggin inhibe la voie BMP. Sa liaison avec BMP4 inhibe d’une part l’activation des gènes épidermiques et d’autre part augmente l’interaction entre le canal TRPC1 et le domaine C-terminal du récepteur BMPRII. Cette interaction qui permet l’activation des canaux TRPC1 et l’entrée de cations (Ca2+ et Na+), induit la dépolarisation de la membrane. Ainsi, le potentiel passe d’environ -55 mV à -20 mV. Cette valeur correspond au seuil d’activation des canaux Cav1.2. L’activation des canaux Cav1.2 permet une entrée de Ca2+ plus importante. Au niveau nucléaire, ce Ca2+ régule directement la transcription de gènes neuraux précoces tels prmt1b et sox2. Prmt1b régule ensuite directement l’expression de zic3. Le mécanisme de régulation fait intervenir le répresseur transcriptionnel KCNIP1. KCNIP1 fait partie de la famille des protéines KCNIP qui constituent la classe E des senseurs de Ca2+. Ce sont des protéines globulaires de 217 à 270 acides aminés qui possèdent un domaine central très conservé contenant 4 motifs de liaison au Ca2+ de type EF-hand (EF-1, EF-2, EF-3 et EF-4). Parmi ces quatre motifs, seul EF-1 n’est pas capable de lier le Ca2+. Les protéines KCNIP contrôlent la transcription des gènes de manière directe. Elles se lient, sous forme de tétramères, à des sites appelés DRE (downstream regulatory element) localisés dans la séquence 5’proximale du gène, en aval de la boîte TATA et en amont du codon START. La séquence GTCA forme le noyau central du site DRE. En absence de Ca2+, le site DRE est occupé et la transcription est réprimée. La liaison du Ca2+ sur les sites EF-hand modifie l’affinité des tétramères KCNIP pour le site DRE, la transcription est alors possible. Dans l’embryon, ce mécanisme original est actif dans l’ectoderme dorsal, destiné à donner du tissu neural.
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.