")
")
Figure 2
Télécharger l'image originale
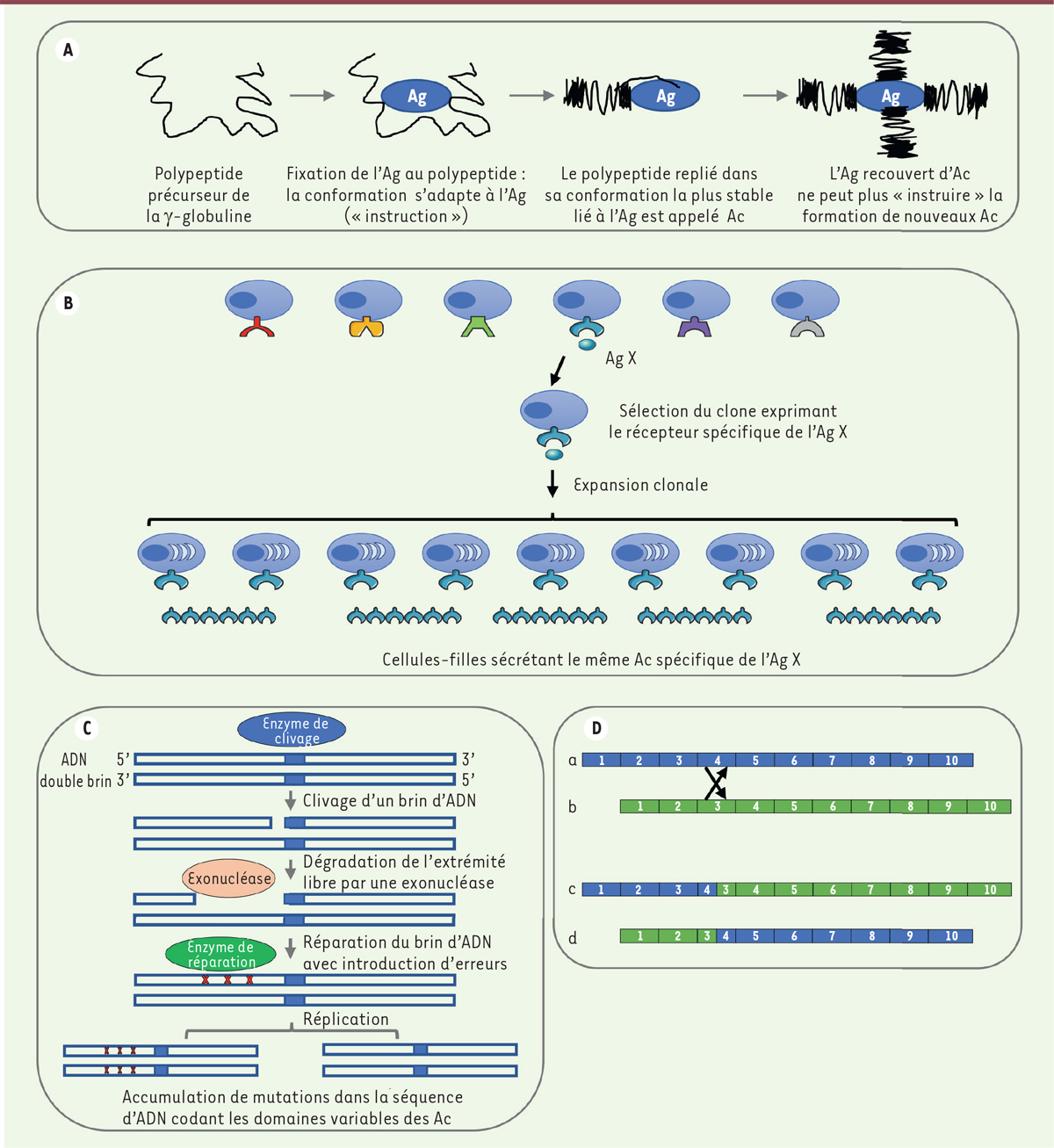
Théories sur l’origine de la diversité de reconnaissance des anticorps. A. Théorie « instructive » de Linus Pauling en 1940 [10]. Dans cette théorie, l’antigène (Ag) est délivré au site de synthèse du polypeptide précurseur de la γ-globuline, dont les extrémités ont une configuration instable et non repliée. L’Ag « attire » des séquences complémentaires de la γ-globuline grâce à des groupements chargés qu’il possède à sa surface, façonnant la conformation de la γ-globuline la plus complémentaire. Cette conformation s’adapte à celle de l’Ag pour rendre l’interaction la plus stable possible, notamment grâce à la flexibilité des extrémités de la γ-globuline. Pour Pauling, la γ-globuline est alors devenue un anticorps (Ac). Dans son modèle, il envisage deux sites de liaison à l’Ag par molécule de γ-globuline. B. Théorie de la sélection clonale de Burnet en 1957 [8]. Cette théorie, confirmée expérimentalement par Nossal et Lederberg l’année suivante (un lymphocyte B = un anticorps), avançait l’hypothèse que chaque lymphocyte B exprime un récepteur spécifique, s’active après fixation de l’Ag correspondant, prolifère, conduisant à une expansion clonale de ce lymphocyte, donnant naissance à des cellules filles (clone) produisant le même anticorps contre cet Ag X. C. Théorie des hypermutations somatiques de Brenner et Milstein en 1966 [35]. Dans cette théorie visionnaire, une enzyme clive un brin de l’ADN du gène codant une chaîne légère, exposant une extrémité 3’. Une exonucléase va ensuite dégrader une partie du brin d’ADN, exposant une partie de la séquence du brin complémentaire. Une ADN polymérase répare le brin d’ADN mais en introduisant des erreurs (délétions et/ou additions de nucléotides et erreurs d’appariements de base) lors de cette réparation, conduisant à l’apparition de mutations somatiques dans la chaîne légère. Le même processus surviendrait pour la chaîne lourde, expliquant ainsi l’origine de la diversité du répertoire des Ac. D. Théorie de la recombinaison somatique de gènes multiples dupliqués d’Edelman et Gally en 1966 [34]. Cette théorie fait l’hypothèse qu’il existe de nombreux gènes qui ont été dupliqués en tandem au cours de l’évolution et qui ont accumulé des mutations ponctuelles, ce qui va favoriser des entrecroisements (« crossing-over ») somatiques entre eux au cours de la maturation du lymphocyte B, permettant d’acquérir une grande diversité de séquences d’acides aminés. Les recombinaisons somatiques ainsi que les erreurs d’appariements de bases sont facilitées par l’homologie des gènes et leur organisation en tandem. Edelman et Gally calculent que ces recombinaisons pourraient permettre de produire jusqu’à 1012 anticorps de spécificités différentes à partir de deux gènes (différents sur vingt nucléotides au minimum) codant les chaines lourdes et deux gènes (différents sur vingt nucléotides au minimum) codant les chaines légères. Dans le schéma, (a et b) montrent l’arrangement de 10 gènes avant recombinaison; (c et d) montrent l’arrangement de ces gènes après recombinaison (adapté de la Figure 2 de [34]).
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.