")
")
| Issue |
Med Sci (Paris)
Volume 34, Number 2, Février 2018
|
|
|---|---|---|
| Page(s) | 112 - 114 | |
| Section | Le Magazine | |
| DOI | https://doi.org/10.1051/medsci/20183402003 | |
| Published online | 16 février 2018 | |
Nouvelles
La sphingosine 1-phosphate, un nouveau régulateur de la mitose
Sphingosine 1-phosphate as a new regulator of mitosis
Institut de pharmacologie et de biologie structurale, université de Toulouse, CNRS, UPS, 205 route de Narbonne, 31077 Toulouse, France
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
La sphingosine 1-phosphate (S1P) est un lipide présent dans une grande diversité d’organismes et qui régule de nombreux processus biologiques [1]. La S1P est issue de la conversion de la sphingosine par les deux isoformes de la sphingosine kinase (SphK1 et SphK2) et, de manière générale, son rôle est opposé à celui de son précurseur, un médiateur de l’arrêt du cycle cellulaire et de l’apoptose [2]. La voie de signalisation de la S1P est fréquemment dérégulée dans le cancer avec une quantité accrue de S1P, conséquence d’une production augmentée et/ ou d’une dégradation réduite [3]. Dans ce contexte, la S1P est impliquée dans la prolifération, la survie, l’invasion et la migration des cellules tumorales ainsi que dans la résistance thérapeutique. Elle participe également aux processus inflammatoires ou angiogéniques associés aux tumeurs [4] ( ).
).
()Voir la Nouvelle de O. Cuvillier, m/s n° 11, novembre 2015, page 964
Cette voie de signalisation représente donc une cible potentielle dans le traitement du cancer [5]. La S1P exerce principalement ses effets biologiques en se liant à l’un des cinq récepteurs S1P1-5 de type RCPG (récepteur couplé aux G-protéines) pour induire une cascade de signalisation paracrine ou autocrine. Ces récepteurs sont exprimés de façon différentielle d’un type cellulaire à l’autre, et selon le contexte physio-pathologique, œuvrant en synergie ou en antagonisme. L’effet biologique observé en réponse à la S1P résulte de l’ensemble de ces actions [6] ().
()Voir la Synthèse de O. Cuvillier, m/s n° 11, novembre 2012, page 951
Alternativement, une signalisation intracellulaire existe, mais les cibles de la S1P sont encore mal définies et même controversées en raison des difficultés techniques inhérentes à l’étude des interactions protéine-lipide et à l’absence d’évidence génétique ou structurale.
La division cellulaire, ou mitose des cellules somatiques, est l’ensemble des évènements de séparation du génome (caryodiérèse) et du corps cellulaire (cytocinèse) permettant la formation des deux cellules filles génétiquement identiques. Au cours de la prométaphase, le fuseau mitotique capture les chromosomes au niveau de leurs centromères/kinétochores afin de les aligner sur la plaque métaphasique. Pour que la ségrégation du matériel génétique soit équitable, chaque chromosome doit établir des connections stables avec les deux pôles du fuseau mitotique en générant un accrochage dit amphitélique. Le point de contrôle du fuseau (spindle assembly checkpoint ou SAC) prévient la séparation des chromatides sœurs tant que tous les chromosomes ne sont pas correctement attachés et alignés. Lorsque tous les chromosomes sont reliés aux deux pôles du fuseau mitotique, le SAC devient inactif et la transition de la métaphase vers l’anaphase est amorcée. L’ensemble de ces mécanismes prévient ainsi les erreurs de ségrégation des chromosomes et la génération d’aneuploïdie, c’est-àdire l’altération du nombre de chromosomes par cellule. À l’instar de l’instabilité chromosomique, l’aneuploïdie est reconnue comme une caractéristique de la plupart des cellules cancéreuses, contribuant à l’initiation et la progression du cancer [7].
Bien que l’effet prolifératif de la S1P ait été rapporté dans pléthore de modèles cellulaires, son rôle dans la régulation du cycle cellulaire et les mécanismes moléculaires sous-jacents associés sont mal compris. Des études ont suggéré un rôle des kinases SphK1 et SphK2 dans la progression de la phase G1 vers S [8]. Quelques travaux proposent un rôle pour la voie SphK1/S1P dans la mitose. L’accumulation de la sphingosine inactiverait la cyclin-dependent kinase 1 (CDK1), une kinase qui s’associe avec la cycline B pour former un complexe contrôlant la transition de la phase G2 vers M et le déroulement de la mitose [9]. L’inhibition de la SphK1 dans un modèle cellulaire de cancer du sein augmente le contenu en ADN, suggérant un défaut de cytocinèse [10]. Collectivement, ces observations suggèrent que la voie de signalisation SphK1/S1P pourrait réguler la division cellulaire.
Nous avons récemment identifié une nouvelle fonction de la signalisation dépendante de la S1P au cours de la mitose [11]. Nous avons observé que la déplétion de la SphK1 causait un arrêt en prométaphase. Inversement, sa surexpression ou son activation induisait une accélération de la mitose. L’inhibition pharmacologique de la SphK1 conduit à un retard dans l’avancée de la mitose similaire à celui observé lors de la déplétion de la SphK1. Ces données suggèrent que la fonction mitotique de la SphK1 dépend de son activité enzymatique et donc de son produit, la S1P. L’utilisation d’un anticorps neutralisant la S1P [12] dans le milieu extracellulaire nous a permis de démontrer que la S1P impactait le déroulement de la mitose de manière autocrine et paracrine. En effet, l’anticorps neutralisant dirigé contre la S1P bloque l’accélération de la mitose induite par l’ajout de S1P dans le milieu ou la surexpression ectopique de la SphK1, dans les modèles cellulaires. Par ailleurs, la déplétion du transporteur membranaire de S1P, Spns2 (spinter homolog 2), bloque la sécrétion de S1P et conduit à un retard dans la mitose. Enfin, dans des cellules humaines HeLa, l’anticorps anti-S1P bloque les perturbations mitotiques induites par l’ajout de milieu conditionné riche en S1P, sécrété par des cellules tumorales prostatiques.
Comme mentionné en introduction, la signalisation extracellulaire de la S1P est relayée par la liaison à l’un des cinq récepteurs (S1P1-5). Par des approches de délétion ou d’inhibition pharmacologique, nous avons identifié le récepteur S1P5 comme la cible centrale de l’effet de la S1P sur la mitose. En accord avec ces résultats, le déroulement de la mitose n’est pas affecté par la S1P dans des fibroblastes embryonnaires de souris (mouse embryonic fibroblast, MEF) issus de souris invalidées pour le gène s1p5. Nous avons pu établir que la signalisation dépendante du S1P5 impliquait la voie PI3K/Akt/Plk1 (phosphatidylinositol-3-kinase/protein kinase B/polo-like kinase 1). La kinase mitotique Plk1 a récemment été identifiée comme une cible directe de la kinase Akt et sa phosphorylation sur la sérine 99 (Ser99), au voisinage du kinétochore, est requise pour une progression normale de la métaphase vers l’anaphase. [13]. L’utilisation d’un système d’expression inductible par la tétracycline (Tet-On) pour générer des lignées HeLa exprimant une protéine PlK1 non-phosphorylable sur sa Ser99, nous a permis de démontrer que l’effet mitotique de la S1P nécessitait cette phosphorylation de la kinase Plk1.
Cette étude met en évidence un rôle insoupçonné de la signalisation de la S1P et du récepteur S1P5 dans le contrôle de la mitose. Comme récemment démontré avec la voie EGF/EGFR (epidermal growth factor/receptor) [14], ces travaux mettent en exergue le rôle du microenvironnement cellulaire dans la coordination de la mitose. De fait, un milieu extracellulaire enrichi en S1P pourrait accélérer la mitose et conduire à des défauts de ségrégation chromosomique (Figure 1), pouvant favoriser l’aneuploïdie. Il sera important d’analyser dans le futur les conséquences d’une ségrégation incorrecte des chromosomes induite par la S1P et d’évaluer le potentiel ciblage thérapeutique de cette voie de signalisation pour inhiber la prolifération tumorale.
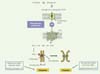 |
Figure 1 Schématisation des voies de sécrétion de S1P et de son action en mitose. La sphingosine 1-phosphate (S1P) est sécrétée grâce au transporteur Spns2, elle interagit avec S1P5, un récepteur lié aux G-protéines qui relaye ses actions. La S1P accélère le déroulement de la mitose et plus particulièrement le déclenchement de l’anaphase. Son action met en jeu la voie PI3K/Akt/Plk1 (phosphatidylinositol-3-kinase/protein kinase B/ polo-like kinase 1) et nécessite la phosphorylation de Plk1 par Akt sur sa Sérine 99. |
Remerciements
Ces travaux ont été soutenus par La Ligue Nationale Contre le Cancer (Comité 31).
Liens d’intérêt
Les auteurs déclarent n’avoir aucun lien d’intérêt concernant les données publiées dans cet article.
Références
- Leong WI, Saba JD. S1P metabolism in cancer and other pathological conditions. Biochimie 2010; 92 : 716-23. [Google Scholar]
- Cuvillier O. Sphingosine in apoptosis signaling. Biochim Biophys Acta 2002; 1585 : 153-62. [CrossRef] [Google Scholar]
- Brizuela L, Ader I, Mazerolles C, et al. First evidence of sphingosine 1-phosphate lyase protein expression and activity downregulation in human neoplasm: implication for resistance to therapeutics in prostate cancer. Mol Cancer Ther 2012; 11 : 1841-51. [Google Scholar]
- Cuvillier O. SphingomabTM, un anticorps antisphingosine 1-phosphate pour inhiber l’hypoxie. Med Sci (Paris) 2015; 31 : 964-7. [Google Scholar]
- Cuvillier O. Downregulating sphingosine kinase-1 for cancer therapy. Expert Opin Ther Targets 2008; 12 : 1009-20. [CrossRef] [Google Scholar]
- Cuvillier O. Récepteurs sphingosine 1-phosphate : de la biologie à la physiopathologie. Med Sci (Paris) 2012; 28 : 951-7. [Google Scholar]
- Torres EM, Williams BR, Amon A. Aneuploidy: cells losing their balance. Genetics 2008; 179 : 737-46. [Google Scholar]
- Igarashi N, Okada T, Hayashi S, et al. Sphingosine kinase 2 is a nuclear protein and inhibits DNA synthesis. J Biol Chem 2003; 278 : 46832-9. [Google Scholar]
- Kim DS, Kim SY, Kleuser B, et al. Sphingosine-1-phosphate inhibits human keratinocyte proliferation via Akt/protein kinase B inactivation. Cell Signal 2004; 16 : 89-95. [Google Scholar]
- Kotelevets N, Fabbro D, Huwiler A, Zangemeister-Wittke U. Targeting sphingosine kinase 1 in carcinoma cells decreases proliferation and survival by compromising PKC activity and cytokinesis. PLoS One 2012; 7 : e39209. [Google Scholar]
- Andrieu G, Ledoux A, Branka S, et al. Sphingosine 1-phosphate signaling through its receptor S1P5 promotes chromosome segregation and mitotic progression. Sci Signal 2017; 10. [Google Scholar]
- Brizuela L, Martin C, Jeannot P, et al. Osteoblastderived sphingosine 1-phosphate to induce proliferation and confer resistance to therapeutics to bone metastasis-derived prostate cancer cells. Mol Oncol 2014; 8 : 1181-95. [Google Scholar]
- Kasahara K, Goto H, Izawa I, et al. PI 3-kinasedependent phosphorylation of Plk1-Ser99 promotes association with 14-3-3gamma and is required for metaphase-anaphase transition. Nat Commun 2013; 4 : 1882. [Google Scholar]
- Mardin BR, Isokane M, Cosenza MR, et al. EGF-induced centrosome separation promotes mitotic progression and cell survival. Dev Cell 2013; 25 : 229-40. [Google Scholar]
© 2018 médecine/sciences – Inserm
Liste des figures
|
Figure 1 Schématisation des voies de sécrétion de S1P et de son action en mitose. La sphingosine 1-phosphate (S1P) est sécrétée grâce au transporteur Spns2, elle interagit avec S1P5, un récepteur lié aux G-protéines qui relaye ses actions. La S1P accélère le déroulement de la mitose et plus particulièrement le déclenchement de l’anaphase. Son action met en jeu la voie PI3K/Akt/Plk1 (phosphatidylinositol-3-kinase/protein kinase B/ polo-like kinase 1) et nécessite la phosphorylation de Plk1 par Akt sur sa Sérine 99. |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.