")
")
| Issue |
Med Sci (Paris)
Volume 29, Number 1, Janvier 2013
|
|
|---|---|---|
| Page(s) | 104 - 106 | |
| Section | Dernières Heures | |
| DOI | https://doi.org/10.1051/medsci/2013291022 | |
| Published online | 25 janvier 2013 | |
Les exosomes
Des transporteurs d’ARN viral qui stimulent l’immunité innée
Exosomes are carriers for immunostimulatory viral RNA
1
Inserm U758, laboratoire de virologie humaine ; École normale supérieure de Lyon ; 46, allée d’Italie, 69364 Lyon Cedex 07, France
2
Université de Lyon, UCB-Lyon1, Lyon, F-69007, France
*
Cette adresse e-mail est protégée contre les robots spammeurs. Vous devez activer le JavaScript pour la visualiser.
Diversité des particules virales produites par les cellules infectées
Les cellules infectées peuvent produire des particules virales qui, infectieuses ou non, revêtent différentes formes et compositions. Les particules virales capables d’infecter de façon productive des cellules cibles comportent, généralement, le génome viral ainsi que les protéines structurales. Le plus souvent, ces particules infectieuses représentent une très faible proportion des particules sécrétées par les cellules infectées. En effet, celles-ci produisent également une grande quantité de particules virales incomplètes qui diffèrent des précédentes par leur composition, notamment en protéines virales structurales, ou par la qualité du génome viral encapsulé. Par exemple, les cellules infectées par le virus de l’hépatite B (VHB) sécrètent de larges quantités de particules non infectieuses, dont la protéine virale antigène de surface (HBsAg), mais sont dépourvues de capside et de génome viral [1]. Par ailleurs, des virus défectifs interférents, composés de génomes viraux tronqués, sont produits dans un large spectre d’infections comme la grippe et certaines infections par flavivirus [2, 3]. Ces génomes tronqués, générés probablement à la suite d’erreurs de la polymérase virale ou par digestion nucléotidique partielle, sont aussi efficacement encapsidés et sécrétés que le génome viral entier [3].
Les particules du virus de l’hépatite C (VHC) sont particulièrement hétérogènes en termes de taille, de densité, de composition et d’infectivité [4]. Cette hétérogénéité est en partie inhérente à la capacité du VHC à s’associer à différents composants des lipoprotéines [4]. Les cellules infectées par le VHC, comme celles qui sont infectées par d’autres virus, produisent 1 particule infectieuse pour environ 100 à 1000 génomes viraux sécrétés [4], ce qui suscite des interrogations quant à la nature et à la fonction des génomes viraux contenus dans ces particules non infectieuses et/ou incomplètes.
Activation de la réponse innée par des exosomes contenant l’ARN viral
Dans une publication récemment parue dans Cell Host Microbe [5], nous avons montré que de larges quantités d’ARN du VHC sont encapsulées dans des vésicules apparentées aux exosomes. Les exosomes sont définis comme des vésicules lipidiques extracellulaires dérivant des corps multivésiculaires (MVB) du compartiment endosomal [6]. Plus spécifiquement, nous avons démontré que ce mode de transfert d’ARN viral par les exosomes ne nécessite pas de protéines virales structurales [5]. Par ailleurs, nos résultats suggèrent que l’ARN du VHC peut ainsi être transféré des cellules infectées aux cellules dendritiques plasmacytoïdes (pDC) par l’intermédiaire de ces exosomes [5] (Figure 1). Les pDC sont des cellules immunitaires capables de produire de grandes quantités de cytokines antivirales, en particulier les interférons de type I (IFNα et IFNβ), médiateurs de la réponse immunitaire innée [7]. En effet, ces derniers induisent l’expression de nombreux effecteurs - codés par les interferon stimulated genes (ISG) - qui bloquent la propagation virale directement en agissant sur les composants viraux, ou indirectement via la mobilisation de la réponse immunitaire adaptative [8]. De manière intéressante, nous avons observé que les exosomes contenant l’ARN viral et produits par des cellules infectées sont capables d’induire une forte production d’IFN par les pDC [5]. Cette activation de la réponse antivirale mettrait en jeu le récepteur Toll-like TLR 7 [5, 9] présent dans les endosomes des pDC et qui reconnaît l’ARN viral simple brin [8]. De plus, nos résultats montrent que l’inhibition de la production d’exosomes par les cellules infectées, en utilisant une approche pharmacologique (les inhibiteurs GW4869 et spiroepoxide), bloque efficacement l’activation des pDC, alors que le relargage de particules infectieuses n’est pas significativement diminué [5]. Ces observations suggèrent donc que le mécanisme de production des exosomes contenant l’ARN viral diffèrerait, du moins en partie, de celui des virus infectieux qui, lui, ne mettrait pas en jeu la voie de sécrétion des exosomes. Par ailleurs, nos données montrent que les particules infectieuses du VHC ne participent pas à l’activation des pDC. En accord avec nos résultats, de récents travaux montrent que la protéine d’enveloppe E2 du VHC interagit avec un récepteur inhibiteur de la réponse innée, BDCA-2, et bloque ainsi l’activation des pDC [10] (Figure 1). L’ensemble de ces observations suggèrent donc une action opposée des exosomes contenant l’ARN du VHC et des particules infectieuses contenant les protéines structurales qui activent et inhibent, respectivement, les pDC.
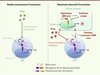 |
Figure 1. Modèles d’activation de la production d’IFN de type I lors d’une infection virale . Selon le modèle précédemment défini, les pDC reconnaissent l’ARN viral via le récepteur endosomal TLR7 après internalisation et/ou infection par certains virus, dont le virus de la grippe, ce qui induit l’activation de la production d’IFN (partie gauche). Cependant, la plupart des virus, notamment le VHC, expriment des protéines qui inhibent la production d’IFN dans les cellules qu’ils infectent. De plus, la liaison de la protéine structurale E2 du VHC aux pDC bloque leur activation. Nous proposons donc un mécanisme alternatif d’activation de la sécrétion d’IFN, selon lequel les pDC ne sont pas stimulées par les virus complets et infectieux mais par des exosomes produits par la cellule infectée, contenant l’ARN viral et dépourvus des protéines structurales (partie droite). |
Nous avons ensuite défini des facteurs de l’hôte impliqués dans ce mécanisme, nouvellement identifié, d’activation de la réponse immunitaire innée. Nous avons démontré que des protéines de la machinerie ESCRT (endosomal sorting complex required for transport) sont nécessaires pour cette activation [5]. Cette machinerie permet l’invagination de vésicules intraluminales au niveau des MVB et participerait à la formation d’exosomes [11]. L’implication des protéines ESCRT dans le mécanisme d’activation des pDC confirme donc le rôle des exosomes comme transporteurs de l’ARN viral immunostimulateur.
De façon intéressante, la protéine cellulaire annexine A2 (ANXA2) est également requise pour la formation du signal activateur des pDC par les cellules infectées [5]. ANXA2 est localisée au niveau des membranes plasmiques et des endosomes, où elle participe à des processus de remodelage membranaire et de trafic intracellulaire [12]. Notamment, ANXA2 favorise la maturation des endosomes précoces en MVB [13]. De plus, elle est recrutée au niveau des sites de réplication du VHC et elle participe à la production de particules infectieuses [14]. L’exacte fonction d’ANXA2 dans le mécanisme d’activation des pDC demeure encore énigmatique et fait l’objet de nos recherches actuelles.
Jusqu’à présent, on pensait que la production d’IFN par les pDC était induite par l’internalisation de virus (par exemple, virus de la grippe). Cependant, l’ensemble de nos résultats révèle l’existence d’un mécanisme alternatif d’activation des pDC via le transfert d’ARN viral immunostimulateur par les exosomes (Figure 1). Il est concevable que ce mécanisme d’activation de la réponse antivirale soit généralisable à d’autres agents infectieux. En faveur de cette hypothèse, on observe des similitudes concernant l’activation des pDC par des virus génétiquement distincts. Ainsi, l’activation des pDC par les cellules qui répliquent un virus de la famille des Togaviridae (le virus de l’encéphalite équine du Venezuela) ne nécessite pas de production de virus complets et infectieux [9]. De plus, comme pour le VHC, la stimulation de la réponse immunitaire innée par cette infection est conditionnée par l’activation de TLR7 [9] et impliquerait donc la reconnaissance de l’ARN viral par les pDC [8].
Rôle fonctionnel des particules virales incomplètes
D’autres fonctions biologiques des particules virales non infectieuses et/ou incomplètes ont été mises en évidence. Par exemple, les particules HBsAg sont capables - comme E2 du VHC - d’inhiber la production d’IFN par les pDC [15]. Par ailleurs, les virus défectifs interférents bloquent la réplication des virus infectieux, notamment en réquisitionnant la polymérase virale et d’autres facteurs pour leur propre réplication [3]. En outre, des particules rétrovirales non infectieuses contenant des génomes mutants générés par l’action de déoxycytidines déaminases - facteurs de restriction comme par exemple APOBEC3 - ont la capacité d’induire une réponse neutralisante, ce qui contribue au contrôle de la charge virale in vivo [16].
La majorité des virus codent pour des protéines qui bloquent la réponse antivirale dans les cellules qu’ils infectent (Figure 1). Notre étude démontre que des particules virales incomplètes, présentant les caractéristiques d’exosomes et incorporant l’ARN viral entier, activent efficacement la production d’IFN par des pDC [5]. Cette réponse n’est pas contrée par des protéines virales, puisque ces pDC ne sont pas infectées de manière productive, c’est-à-dire qu’elles ne répliquent pas le virus [17]. Ce mécanisme alternatif de détection de l’infection virale par les cellules immunitaires avoisinantes permet donc à l’hôte de réagir contre le virus, alors même que la réponse innée est bloquée dans les cellules infectées (Figure 1).
Ces études dévoilent que des particules virales non infectieuses et/ou incomplètes jouent un rôle clé dans la propagation virale, directement ou indirectement via la modulation de la réponse immunitaire. En fonction du mécanisme de détection de ces particules défectueuses par l’hôte et, par conséquent, de la voie immunitaire qui est modulée, l’impact sur la propagation virale peut être négatif ou positif. La poursuite de ces études offre un nouvel angle pour le décryptage de la complexité des relations hôte-pathogène.
Liens d’intérêt
Les auteurs déclarent n’avoir aucun lien d’intérêt concernant les données publiées dans cet article.
Références
- Howard CR. The biology of hepadnaviruses. J Gen Virol 1986 ; 67 : 1215–1235. [CrossRef] [PubMed] [Google Scholar]
- Pijlman GP, Funk A, Kondratieva N, et al. A highly structured, nuclease-resistant, noncoding RNA produced by flaviviruses is required for pathogenicity. Cell Host Microbe 2008 ; 4 : 579–591. [CrossRef] [PubMed] [Google Scholar]
- Marriott AC, Dimmock NJ. Defective interfering viruses and their potential as antiviral agents. Rev Med Virol 2010 ; 20 : 51–62. [CrossRef] [PubMed] [Google Scholar]
- Bartenschlager R, Penin F, Lohmann V, Andre P. Assembly of infectious hepatitis C virus particles. Trends Microbiol 2011 ; 19 : 95–103. [Google Scholar]
- Dreux M, Garaigorta U, Boyd B, et al. Short-range exosomal transfer of viral RNA from infected cells to plasmacytoid dendritic cells triggers innate immunity. Cell Host Microbe 2012 ; 12 : 558–570. [CrossRef] [PubMed] [Google Scholar]
- Thery C, Zitvogel L, Amigorena S. Exosomes: composition, biogenesis and function. Nat Rev Immunol 2002 ; 2 : 569–579. [PubMed] [Google Scholar]
- Reizis B, Bunin A, Ghosh HS, et al. Plasmacytoid dendritic cells: recent progress and open questions. Annu Rev Immunol 2011 ; 29 : 163–183. [CrossRef] [PubMed] [Google Scholar]
- Kawai T, Akira S. The roles of TLRs, RLRs and NLRs in pathogen recognition. Int Immunol 2009 ; 21 : 317–337. [CrossRef] [PubMed] [Google Scholar]
- Takahashi K, Asabe S, Wieland S, et al. Plasmacytoid dendritic cells sense hepatitis C virus-infected cells, produce interferon, and inhibit infection. Proc Natl Acad Sci USA 2010 ; 107 : 7431–7436. [CrossRef] [Google Scholar]
- Florentin J, Aouar B, Dental C, et al. HCV glycoprotein E2 is a novel BDCA-2 ligand and acts as an inhibitor of IFN production by plasmacytoid dendritic cells. Blood 2012 ; 120 : 4544–4551. [CrossRef] [PubMed] [Google Scholar]
- Baietti MF, Zhang Z, Mortier E, et al. Syndecan-syntenin-ALIX regulates the biogenesis of exosomes. Nat Cell Biol 2012 ; 14 : 677–685. [CrossRef] [PubMed] [Google Scholar]
- Gerke V, Creutz CE, Moss SE. Annexins: linking Ca2+ signalling to membrane dynamics. Nat Rev Mol Cell Biol 2005 ; 6 : 449–461. [CrossRef] [PubMed] [Google Scholar]
- Mayran N, Parton RG, Gruenberg J. Annexin II regulates multivesicular endosome biogenesis in the degradation pathway of animal cells. EMBO J 2003 ; 22 : 3242–3253. [CrossRef] [PubMed] [Google Scholar]
- Backes P, Quinkert D, Reiss S, et al. Role of annexin A2 in the production of infectious hepatitis C virus particles. J Virol 2010 ; 84 : 5775–5789. [CrossRef] [PubMed] [Google Scholar]
- Xu Y, Hu Y, Shi B, et al. HBsAg inhibits TLR9-mediated activation and IFN-alpha production in plasmacytoid dendritic cells. Mol Immunol 2009 ; 46 : 2640–2646. [CrossRef] [PubMed] [Google Scholar]
- Smith DS, Guo K, Barrett BS, et al. Noninfectious retrovirus particles drive the APOBEC3/Rfv3 dependent neutralizing antibody response. PLoS Pathog 2011 ; 7 : e1002284. [CrossRef] [PubMed] [Google Scholar]
- Decalf J, Fernandes S, Longman R, et al. Plasmacytoid dendritic cells initiate a complex chemokine and cytokine network and are a viable drug target in chronic HCV patients. J Exp Med 2007 ; 204 : 2423–2437. [CrossRef] [PubMed] [Google Scholar]
© 2013 médecine/sciences – Inserm / SRMS
Liste des figures
|
Figure 1. Modèles d’activation de la production d’IFN de type I lors d’une infection virale . Selon le modèle précédemment défini, les pDC reconnaissent l’ARN viral via le récepteur endosomal TLR7 après internalisation et/ou infection par certains virus, dont le virus de la grippe, ce qui induit l’activation de la production d’IFN (partie gauche). Cependant, la plupart des virus, notamment le VHC, expriment des protéines qui inhibent la production d’IFN dans les cellules qu’ils infectent. De plus, la liaison de la protéine structurale E2 du VHC aux pDC bloque leur activation. Nous proposons donc un mécanisme alternatif d’activation de la sécrétion d’IFN, selon lequel les pDC ne sont pas stimulées par les virus complets et infectieux mais par des exosomes produits par la cellule infectée, contenant l’ARN viral et dépourvus des protéines structurales (partie droite). |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.