")
")
| Issue |
Med Sci (Paris)
Volume 28, Number 4, Avril 2012
|
|
|---|---|---|
| Page(s) | 368 - 370 | |
| Section | Nouvelles | |
| DOI | https://doi.org/10.1051/medsci/2012284012 | |
| Published online | 25 April 2012 | |
Quel prix à payer pour notre gros cerveau ?
Remise en cause de « la théorie des tissus coûteux »
The expensive tissue hypothesis revisited
Anthropologisches Institut und Museum, Universität Zürich-Irchel, Winterthurerstrasse 190, 8057 Zurich, Suisse
*
This email address is being protected from spambots. You need JavaScript enabled to view it.
Le cerveau est un organe dont le fonctionnement est très coûteux sur le plan énergétique, et qui, contrairement à d’autres tissus, ne peut, même pour un temps très court, se passer d’apports nutritifs [1]. Le cerveau humain, par exemple, ne représente que 2 % du poids du corps, mais est à lui seul responsable de 20 % de la dépense énergétique chez un adulte et de plus de 60 % chez un nouveau-né [2]. Comment l’organisme a-t-il trouvé le surplus d’énergie nécessaire à l’augmentation de la taille du cerveau au cours de l’évolution [1] ? The expensive tissue hypothesis - ou « théorie des tissus coûteux » - proposée par Aiello et Wheeler en 1995 est un des concepts les plus fréquemment avancés pour expliquer ce processus [3].
Le cerveau est un organe dont le fonctionnement est très coûteux sur le plan énergétique, et qui, contrairement à d’autres tissus, ne peut, même pour un temps très court, se passer d’apports nutritifs [1]. Le cerveau humain, par exemple, ne représente que 2 % du poids du corps, mais est à lui seul responsable de 20 % de la dépense énergétique chez un adulte et de plus de 60 % chez un nouveau-né [2]. Comment l’organisme a-t-il trouvé le surplus d’énergie nécessaire à l’augmentation de la taille du cerveau au cours de l’évolution [1] ? The expensive tissue hypothesis - ou « théorie des tissus coûteux » - proposée par Aiello et Wheeler en 1995 est un des concepts les plus fréquemment avancés pour expliquer ce processus [3].
Faut-il réduire la taille du tube digestif pour permettre la croissance du cerveau ?
Cette « théorie des tissus coûteux » avait pour objectif de proposer une explication à l’expansion particulièrement impressionnante du cerveau dans le lignage humain par comparaison aux autres mammifères. Elle stipulait que face à un taux de base constant du métabolisme corporel, l’énergie nécessaire à l’élargissement du cerveau humain devait nécessairement se faire aux dépens d’un allègement de la dépense énergétique (et de la taille) d’autres organes. Mais encore fallait-il que cette réduction ne mette pas en péril la fonction de ces organes. Selon Aiello et Wheeler [3], un seul organe pouvait supporter ces contraintes : le tube digestif. Il était en effet concevable qu’un régime de meilleure qualité (consommation de viande, de racines, cuisson des aliments) puisse permettre une réduction de la masse du tube digestif, épargnant ainsi de l’énergie qui pourrait être dépensée pour le développement du cerveau. À l’appui de leur hypothèse, les auteurs établirent chez 18 primates anthropoïdes (singes, grands singes et hommes) dont homo sapiens, l’existence d’une corrélation négative entre la masse du cerveau et celle du tube digestif. Pourtant, dans un article ultérieur [4], Aiello revenait sur ses conclusions car les données morphologiques qui avaient servi à vérifier l’hypothèse étaient entâchées d’imprécisions : en effet, elles ne provenaient pas d’une seule source puisqu’il s’agissait d’une compilation de données morphologiques collectées avec des méthodologies très diverses. Quoiqu’il en soit, la « théorie des tissus coûteux » a été adoptée car elle était séduisante intuitivement, et les études qui ont tenté par la suite d’en tester la validité dans d’autres groupes de mammifères se sont heurtées au manque de données morphologiques solides.
La « théorie des tissus coûteux » est contestable
Dans une étude récemment publiée dans Nature [5], nous avons évalué cette « théorie des tissus coûteux » chez les mammifères dont les primates, avec une méthodologie rigoureuse. Dans ce but, nous avons accumulé de nouvelles données morphologiques directement à partir de dissections. Au cours de celles-ci, nous avons analysé huit organes (cerveau, poumons, cœur, reins, rate, foie, estomac et intestin) et le tissu adipeux. Nous avons travaillé sur 100 espèces de mammifères, au sein desquelles nous avons isolé un échantillon de 23 espèces de primates. Nous avons analysé la corrélation entre le cerveau et les autres organes en utilisant des analyses phylogénétiques et en tenant compte de la masse corporelle indépendamment du tissu adipeux. Nous n’avons pas tenu compte du tissu graisseux dans notre appréciation de la taille du corps, contrairement à ce qui est fait habituellement, afin de minimiser les variations importantes dues aux dépôts adipeux.
Contrairement aux prédictions de la « théorie des tissus coûteux », nous n’avons pas trouvé de corrélation négative entre la masse du cerveau et celle du tube digestif, que ce soit chez les mammifères en général ou dans le sous-groupe de primates (Figure 1). De plus, une telle corrélation négative n’existait pas non plus entre la masse du cerveau et celle des autres tissus « énergivores » ou entre la masse du cerveau et la somme des masses de ces tissus viscéraux. Nos résultats contredisent donc the expensive tissue hypothesis qui veut que l’expansion du cerveau se soit faite aux dépens de celle du tube digestif. Nous proposons que ce « troc » se fasse plutôt entre le cerveau et le tissu adipeux : en effet, lors de notre recherche de corrélations entre le cerveau et d’autres tissus moins énergivores mais plus abondants, nous avons observé une relation inverse entre la masse du cerveau et celle du tissu adipeux chez les mammifères mais pas chez les primates. Toutefois, il est possible que l’absence de corrélation négative chez ces derniers soit due au fait que nos spécimens provenaient tous d’individus maintenus en captivité.
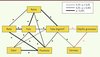 |
Figure 1. Corrélations entre les masses des viscères, du cerveau et des dépôts de tissu adipeux chez les mammifères. La seule corrélation négative qui existe avec la masse du cerveau est celle des dépôts adipeux (trait rouge), alors que la masse de tous les organes viscéraux a tendance à être corrélée positivement (adapté de [5]). |
Cerveau et tissu adipeux : rivaux ou compères ?
Si le tissu adipeux est « bon marché » sur le plan métabolique, il peut malgré tout avoir un coût parce qu’il accroît le poids du corps. De fait, cette entrave à la locomotion peut aggraver le risque de prédation et ses conséquences fatales. A contrario, des réserves graisseuses peuvent être bénéfiques en cas de jeûne. Un cerveau très développé pourrait aussi s’avérer bénéfique en cas de jeûne : on peut imaginer qu’il offre à l’individu les facultés cognitives qui lui permettraient de trouver de nouvelles ressources alimentaires et donc de combattre la disette. Ces deux tissus, cerveau et tissu adipeux, pourraient donc agir de façon complémentaire pour diminuer les conséquences d’une baisse des apports nutritifs. Deux cas de figures se présentent chez les mammifères (notre étude ne s’applique qu’à cette espèce) : chez les animaux qui se déplacent en grimpant ou en volant, ce qui implique une dépense énergétique importante en cas de poids corporel élevé, la corrélation inverse cerveau - tissu adipeux se vérifiera. Chez les mammifères aquatiques ou les grands bipèdes, dont les humains, le coût de déplacement est faible comparativement ; ces espèces mammifères pourraient à la fois profiter de l’intelligence que procure un cerveau de taille importante et d’une accumulation de graisse.
Le consensus actuel : the expensive brain framework ou le « paradigme du cerveau coûteux »
Ces résultats récents ont contribué à mieux appréhender la question du « coût » de l’évolution du cerveau. Ils ont permis, au même titre que d’autres données récentes sur les autres facteurs influençant l’encéphalisation, de construire un schéma général de l’évolution de la taille du cerveau chez les mammifères. Ce schéma général - que plusieurs études comparatives ont contribué à façonner - a été proposé il y a quelques années sous le terme de expensive brain framework que l’on peut traduire par le « paradigme du cerveau coûteux » [6] (Figure 2). Ce cadre propose deux voies - non mutuellement exclusives - qui auraient facilité l’expansion du cerveau à partir d’un stade ancestral : apport énergétique additionnel et détournement de sources d’énergie venant d’autres fonctions.
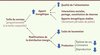 |
Figure 2. The expensive brain framework. L’expansion du cerveau est possible à condition que l’apport énergétique soit stabilisé ou que la dépense énergétique dévolue à d’autres fonctions soit réduite (adapté de [5]). |
Ajouter ou stabiliser l’énergie disponible pour le cerveau peut se faire via l’accroissement du métabolisme ou la réduction de la variation des apports énergétiques. Celle-ci peut être obtenue via l’amélioration du régime, le développement des interactions sociales et la réduction des fluctuations des apports énergétiques (utilisation d’outils, d’instruments de cuisine, etc.). Dans la lignée Homo, la consommation de viande cuite a pu y contribuer significativement. Dans l’espèce humaine, la prise de repas en commun, le partage de nourriture, l’accumulation de provisions ont certainement facilité la constance de l’apport énergétique, en particulier celui des femmes en âge de procréer et de leurs petits très dépendants, favorisant l’élargissement du cerveau. Enfin, les capacités cognitives qu’offre ce « gros » cerveau ont permis de réduire les fluctuations dans l’apport d’énergie au cours du temps et notamment durant les saisons où cet apport est limité, comme le font également d’autres primates et les oiseaux.
La seconde voie consiste à réduire l’énergie dévolue à d’autres fonctions énergivores. Si ceci ne s’applique pas à d’autres organes comme le suggérait the expensive tissue hypothesis que nous avons remise en cause, en revanche la contribution de la locomotion a été importante. L’acquisition de moyens de déplacement plus efficaces, comme le bipédisme, a certainement contribué à l’expansion du cerveau, du moins dans le cas de l’espèce humaine. Ce nouveau mode de locomotion est moins gourmand en énergie que celui des quadrupèdes ou des animaux grimpants, et réduit les conséquences d’une augmentation du poids du corps que provoquent les dépôts de graisse. L’acquisition d’un cerveau de taille importante a pu aussi être favorisée par une réduction des coûts de production, non seulement lors du processus de reproduction, mais aussi en raison de la diminution du rythme de nos vies. Cela n’a pas empêché l’accroissement du taux des naissances dans nos espèces, certainement influencé par le rôle qu’y joue l’assistance des membres de la famille lors des naissances et le partage des ressources.
Nous proposons que, dans notre espèce, la qualité accrue de notre régime alimentaire, mais aussi les interactions sociales - le partage des ressources et l’assistance notamment aux femelles -, l’importance des fonctions cognitives permettant de surmonter les conditions défavorables, une sédentarité accrue et une diminution de l’énergie allouée à la production, ont contribué ensemble au développement extraordinaire du cerveau.
Conflit d’intérêts
L’auteur déclare n’avoir aucun conflit d’intérêts concernant les données publiées dans cet article.
Remerciements
Je remercie Laure Coulombel d’avoir manifesté un si grand intérêt pour nos travaux et d’avoir assuré la traduction française de notre article.
Traduction de l’anglais par Laure Coulombel. Merci à Thibaud Gruber de sa relecture.
Références
- Mink JW, Blumenschine RJ, Adams DB. Ratio of central nervous system to body metabolism in vertebrates: its constancy and functional basis. Am J Physiol 1981 ; 241: R203–R212. [PubMed] [Google Scholar]
- Holliday MA. Human growth: a comprehensive treatise, vol. 2. In : Falkner F, Tanner JM, eds. New York : Plenum Press, 1986 : 101–107. [Google Scholar]
- Aiello LC, Wheeler P. The expensive-tissue hypothesis: the brain and the digestive-system in human and primate evolution. Curr Anthropol 1995 ; 36 : 199–221. [CrossRef] [Google Scholar]
- Aiello LC, Bates N, Joffe T. Evolutionary anatomy of the primate cerebral cortex. In : Falk D, Gibson KR, eds. Cambridge : Cambridge University Press, 2001 : 57–78. [CrossRef] [Google Scholar]
- Navarrete A, van Schaik CP, Isler K. Energetics and the evolution of human brain size. Nature 2011 ; 480 : 91–93. [CrossRef] [PubMed] [Google Scholar]
- Isler K, van Schaik CP. The expensive brain: a framework for explaining evolutionary changes in brain size. J Hum Evol 2009 ; 57 : 392–400. [CrossRef] [PubMed] [Google Scholar]
© 2012 médecine/sciences – Inserm / SRMS
Liste des figures
|
Figure 1. Corrélations entre les masses des viscères, du cerveau et des dépôts de tissu adipeux chez les mammifères. La seule corrélation négative qui existe avec la masse du cerveau est celle des dépôts adipeux (trait rouge), alors que la masse de tous les organes viscéraux a tendance à être corrélée positivement (adapté de [5]). |
| Dans le texte | |
|
Figure 2. The expensive brain framework. L’expansion du cerveau est possible à condition que l’apport énergétique soit stabilisé ou que la dépense énergétique dévolue à d’autres fonctions soit réduite (adapté de [5]). |
| Dans le texte | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.